

Suggestive evidence for Darwinian Selection against asparagine-linked glycans of Plasmodium falciparum and Toxoplasma gondii
- PMID: 19783771
- PMCID: PMC2823003
- DOI: 10.1128/EC.00197-09
Suggestive evidence for Darwinian Selection against asparagine-linked glycans of Plasmodium falciparum and Toxoplasma gondii
Abstract
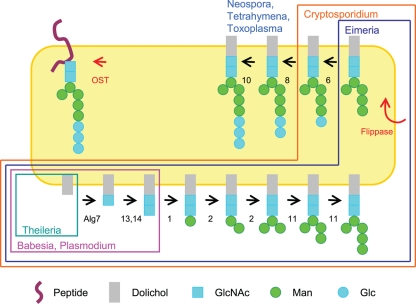
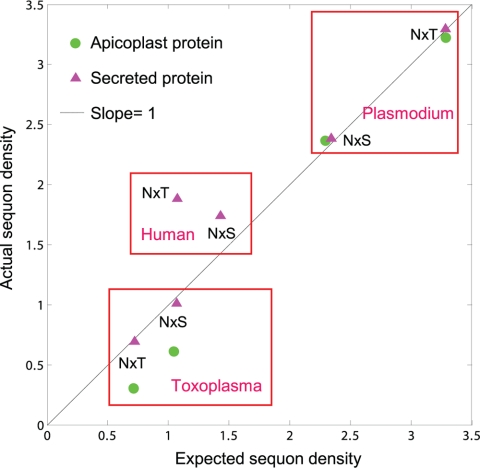
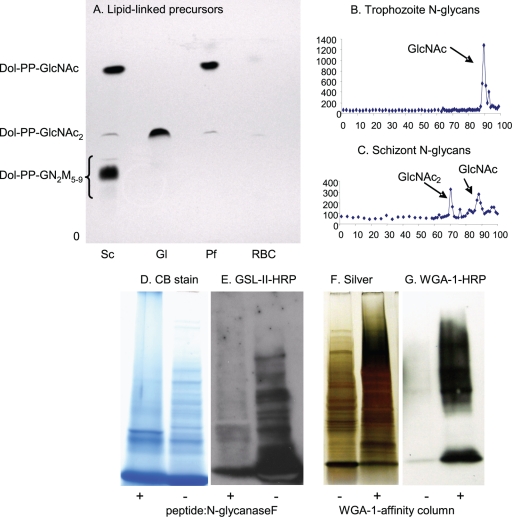
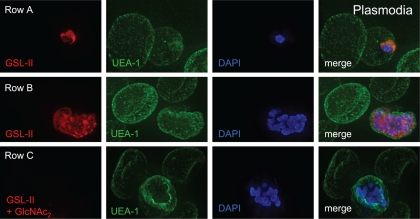
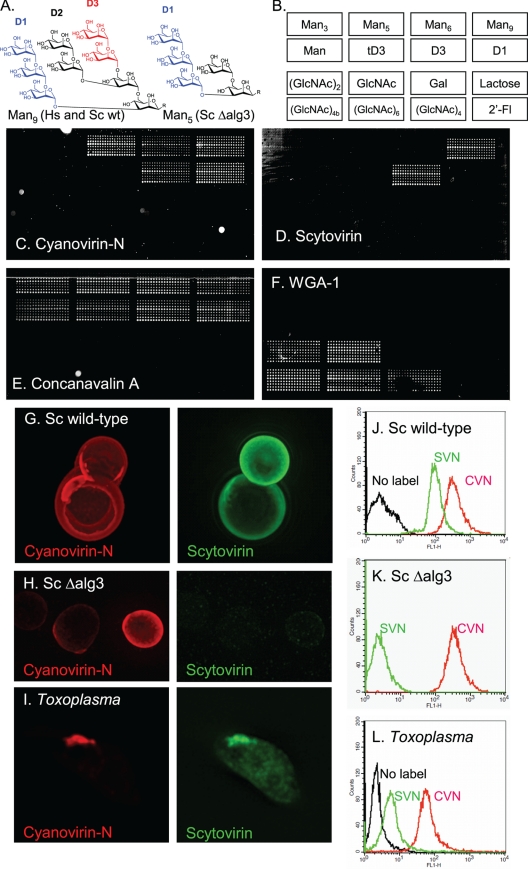
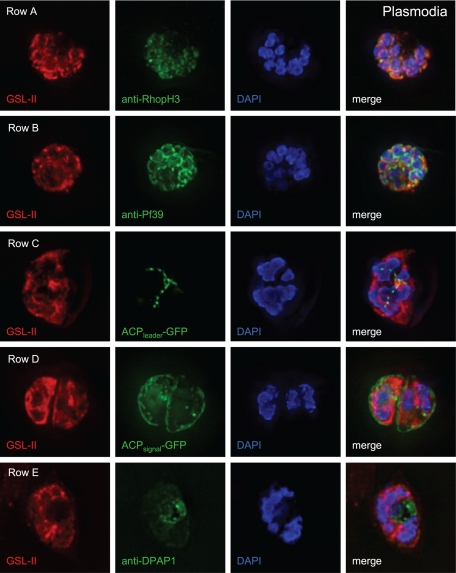
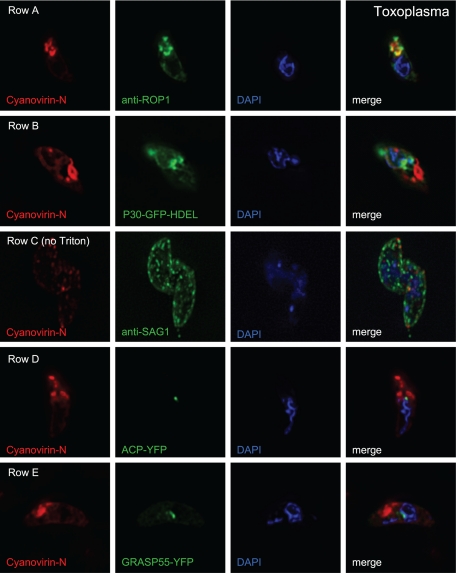
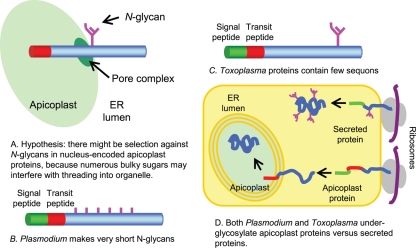
We are interested in asparagine-linked glycans (N-glycans) of Plasmodium falciparum and Toxoplasma gondii, because their N-glycan structures have been controversial and because we hypothesize that there might be selection against N-glycans in nucleus-encoded proteins that must pass through the endoplasmic reticulum (ER) prior to threading into the apicoplast. In support of our hypothesis, we observed the following. First, in protists with apicoplasts, there is extensive secondary loss of Alg enzymes that make lipid-linked precursors to N-glycans. Theileria makes no N-glycans, and Plasmodium makes a severely truncated N-glycan precursor composed of one or two GlcNAc residues. Second, secreted proteins of Toxoplasma, which uses its own 10-sugar precursor (Glc(3)Man(5)GlcNAc(2)) and the host 14-sugar precursor (Glc(3)Man(9)GlcNAc(2)) to make N-glycans, have very few sites for N glycosylation, and there is additional selection against N-glycan sites in its apicoplast-targeted proteins. Third, while the GlcNAc-binding Griffonia simplicifolia lectin II labels ER, rhoptries, and surface of plasmodia, there is no apicoplast labeling. Similarly, the antiretroviral lectin cyanovirin-N, which binds to N-glycans of Toxoplasma, labels ER and rhoptries, but there is no apicoplast labeling. We conclude that possible selection against N-glycans in protists with apicoplasts occurs by eliminating N-glycans (Theileria), reducing their length (Plasmodium), or reducing the number of N-glycan sites (Toxoplasma). In addition, occupation of N-glycan sites is markedly reduced in apicoplast proteins versus some secretory proteins in both Plasmodium and Toxoplasma.
Figures
References
-
- Abrahamsen M. S., Templeton T. J., Enomoto S., Abrahante J. E., Zhu G., et al. 2004. Complete genome sequence of the apicomplexan, Cryptosporidium parvum. Science 304:441–445 - PubMed
-
- Adams E. W., Ratner D. M., Bokesch H. R., McMahon J. B., O'Keefe B. R., Seeberger P. H. 2004. Oligosaccharide and glycoprotein microarrays as tools in HIV glycobiology; glycan-dependent gp120/protein interactions. Chem. Biol. 11:875–881 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
