

The single polypeptide restriction-modification enzyme LlaGI is a self-contained molecular motor that translocates DNA loops
- PMID: 19783815
- PMCID: PMC2790907
- DOI: 10.1093/nar/gkp794
The single polypeptide restriction-modification enzyme LlaGI is a self-contained molecular motor that translocates DNA loops
Abstract
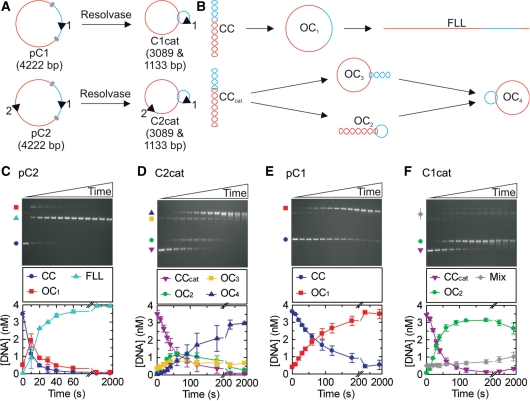
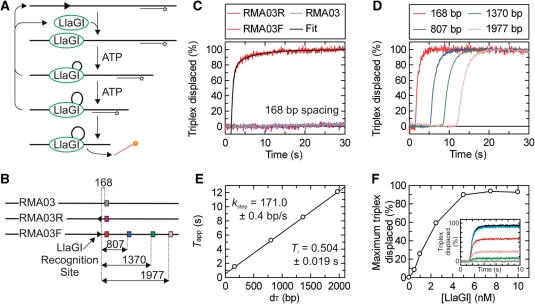
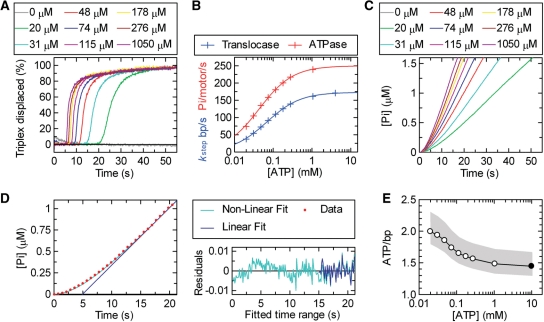
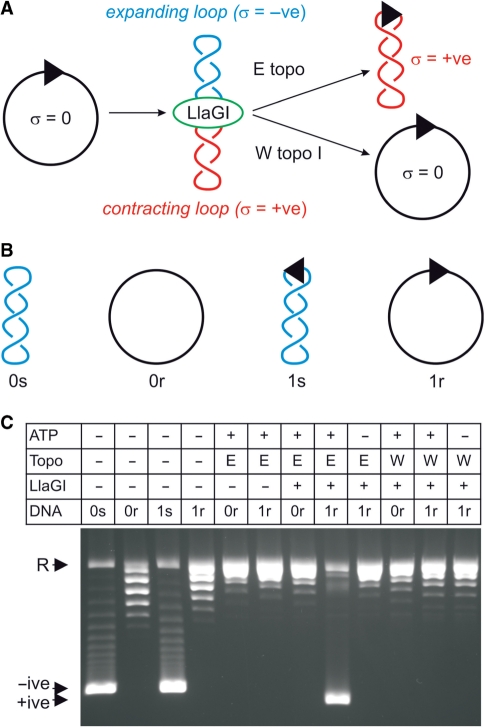
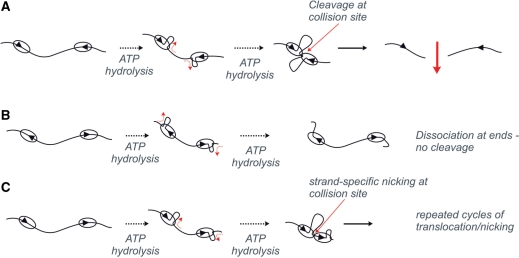
To cleave DNA, the single polypeptide restriction-modification enzyme LlaGI must communicate between a pair of indirectly repeated recognition sites. We demonstrate that this communication occurs by a 1-dimensional route, namely unidirectional dsDNA loop translocation rightward of the specific recognition sequence 5'-CTnGAyG-3' as written (where n is either A, G, C or T and y is either C or T). Motion across thousands of base pairs is catalysed by the helicase domain and requires the hydrolysis of 1.5-2 ATP per base pair. DNA loop extrusion is accompanied by changes in DNA twist consistent with the motor following the helical pitch of the polynucleotide track. LlaGI is therefore an example of a polypeptide that is a completely self-contained, multi-functional molecular machine.
Figures
References
-
- Winkler FK, Prote AE. Structure and function of EcoRV endonuclease in restriction enzymes. In: Pingound A, editor. Nucleic Acids and Molecular Biology. Vol. 14. Germany: Springer; 2004. pp. 179–210.
-
- Bourniquel AA, Bickle TA. Complex restriction enzymes: NTP-driven molecular motors. Biochimie. 2002;84:1047–1059. - PubMed
-
- Halford SE, Welsh AJ, Szczelkun MD. Enzyme-mediated DNA looping. Annu. Rev. Biophys. Biomol. Struct. 2004;33:1–24. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
