

The role of zinc in the modulation of neuronal proliferation and apoptosis
- PMID: 19784710
- PMCID: PMC2797425
- DOI: 10.1007/s12640-009-9067-4
The role of zinc in the modulation of neuronal proliferation and apoptosis
Abstract
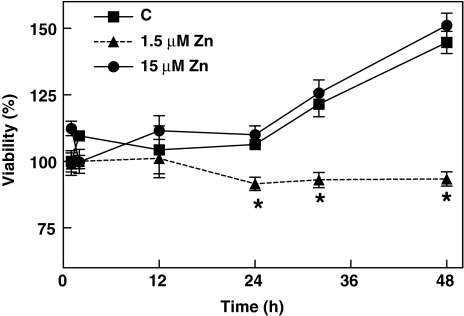
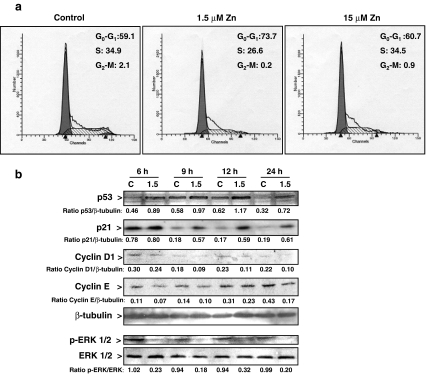
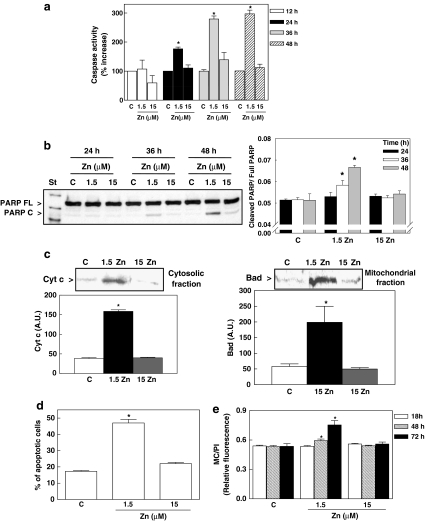
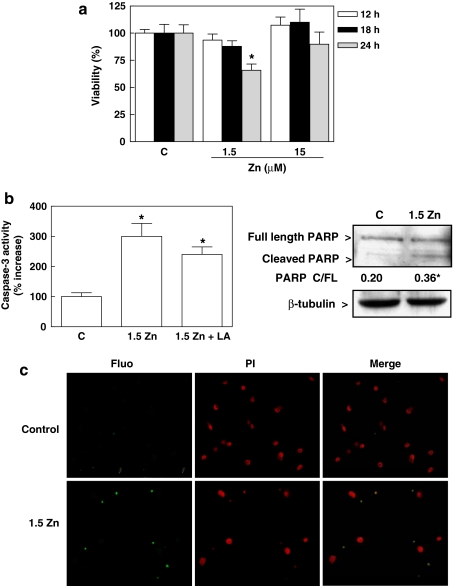
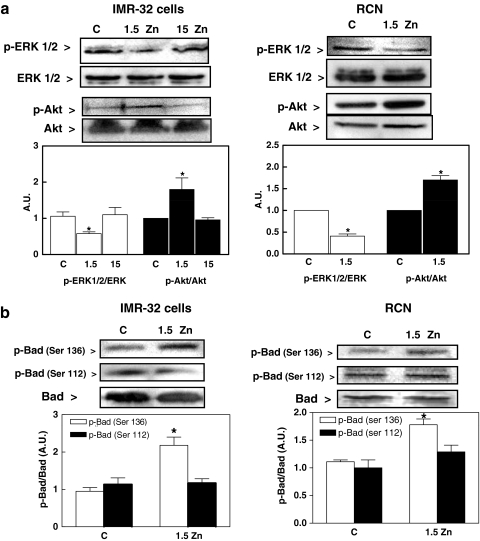
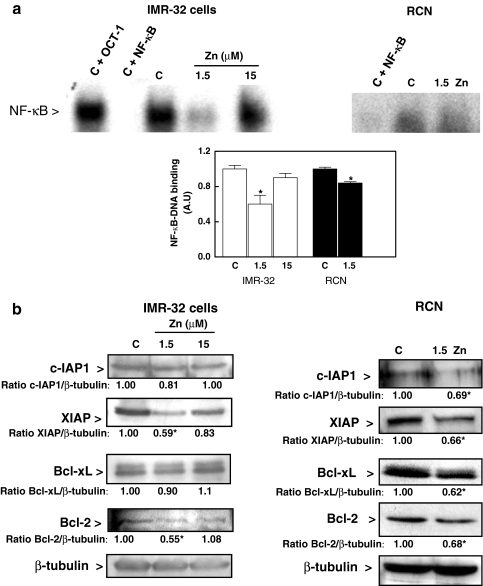
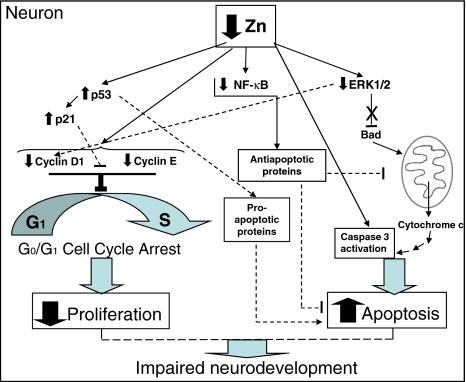
Although a requirement of zinc (Zn) for normal brain development is well documented, the extent to which Zn can modulate neuronal proliferation and apoptosis is not clear. Thus, we investigated the role of Zn in the regulation of these two critical events. A low Zn availability leads to decreased cell viability in human neuroblastoma IMR-32 cells and primary cultures of rat cortical neurons. This occurs in part as a consequence of decreased cell proliferation and increased apoptotic cell death. In IMR-32 cells, Zn deficiency led to the inhibition of cell proliferation through the arrest of the cell cycle at the G0/G1 phase. Zn deficiency induced apoptosis in both proliferating and quiescent neuronal cells via the intrinsic apoptotic pathway. Reductions in cellular Zn triggered a translocation of the pro-apoptotic protein Bad to the mitochondria, cytochrome c release, and caspase-3 activation. Apoptosis is the resultant of the inhibition of the prosurvival extracellular-signal-regulated kinase, the inhibition of nuclear factor-kappa B, and associated decreased expression of antiapoptotic proteins, and to a direct activation of caspase-3. A deficit of Zn during critical developmental periods can have persistent effects on brain function secondary to a deregulation of neuronal proliferation and apoptosis.
Figures
References
-
- Banker G, Goslin K (eds) (1998) Culturing nerve cells, 2nd edn. MIT, Cambridge, MA, pp 11–36
-
- Bentley ME, Caulfield LE, Ram M, Santizo MC, Hurtado E, Rivera JA, Ruel MT, Brown KH. Zinc supplementation affects the activity patterns of rural Guatemalan infants. J Nutr. 1997;127:1333–1338. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
