

Regulation of the epithelial sodium channel [ENaC] in kidneys of salt-sensitive Dahl rats: insights on alternative splicing
- PMID: 19785774
- PMCID: PMC2761857
- DOI: 10.1186/1755-7682-2-28
Regulation of the epithelial sodium channel [ENaC] in kidneys of salt-sensitive Dahl rats: insights on alternative splicing
Abstract
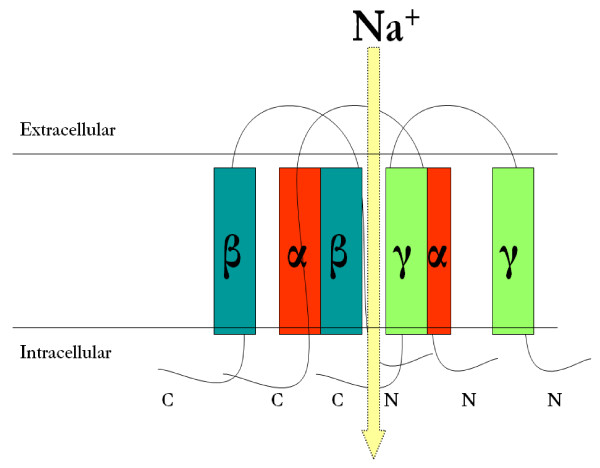
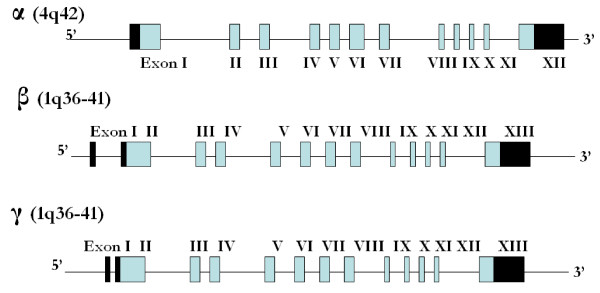
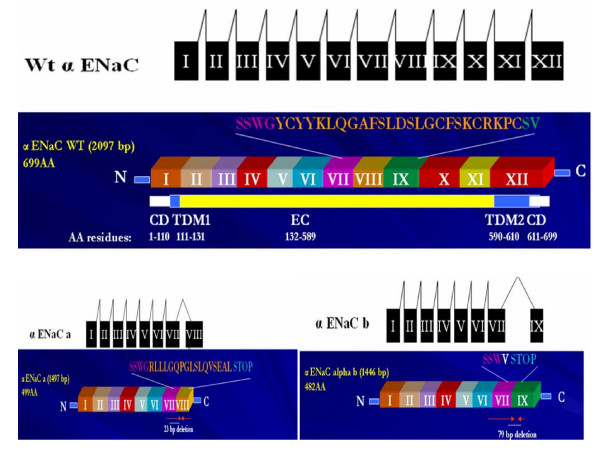
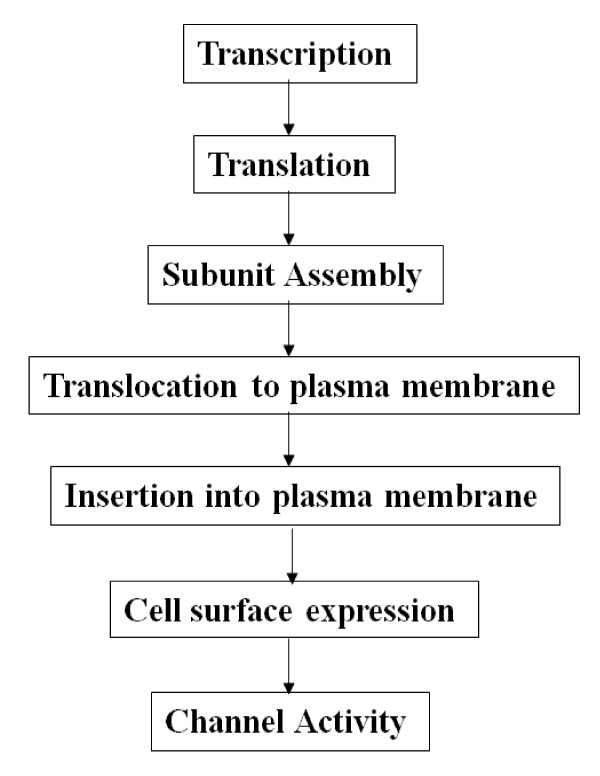
The epithelial sodium channel [ENaC] is critical for the maintenance of sodium balance, extracellular fluid volume and long term blood pressure control. Monogenic disorders causing ENaC hyperactivity have led to a severe form of hereditary hypertension in humans, known as Liddle's syndrome. Similarly, in animal models, ENaC hyperactivity has been well documented in kidneys of salt-sensitive [S] Dahl rats [a genetic model of salt-sensitive hypertension] versus their normotensive control [Dahl salt-resistant [R] rats]. The purpose of the present review is to highlight the differential regulation of ENaC in kidneys of Dahl S versus R rats. A systematic overview of the putative role of alternative splicing of the main alpha subunit of ENaC [alpha ENaC] in modulating ENaC expression in kidneys of Dahl rats will be discussed. Finally, a better understanding of the meaningful contribution of ENaC in the pathogenesis of salt-sensitive hypertension will be achieved upon completion of this review.
Figures

References
-
- Hemmelgarn BR, McAlister FA, Grover S, Myers MG, McKay DW, Bolli P, Abbott C, Schiffrin EL, Honos G, Burgess E, Mann K, Wilson T, Penner B, Tremblay G, Milot A, Chockalingam A, Touyz RM, Tobe SW, Canadian Hypertension Education Program The 2006 Canadian Hypertension Education Program recommendations for the management of hypertension: Part I--Blood pressure measurement, diagnosis and assessment of risk. Can J Cardiol. 2006;22:573–581. - PMC - PubMed
-
- Weinberger MH, Fineberg NS, Fineberg SE, Weinberger M. Salt sensitivity, pulse pressure, and death in normal and hypertensive humans. Hypertension. 2001;37:429–432. - PubMed
LinkOut - more resources
Full Text Sources