

A search for small noncoding RNAs in Staphylococcus aureus reveals a conserved sequence motif for regulation
- PMID: 19786493
- PMCID: PMC2790875
- DOI: 10.1093/nar/gkp668
A search for small noncoding RNAs in Staphylococcus aureus reveals a conserved sequence motif for regulation
Abstract
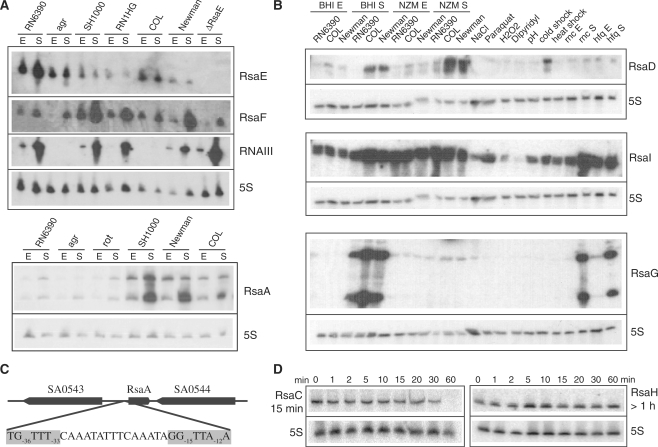
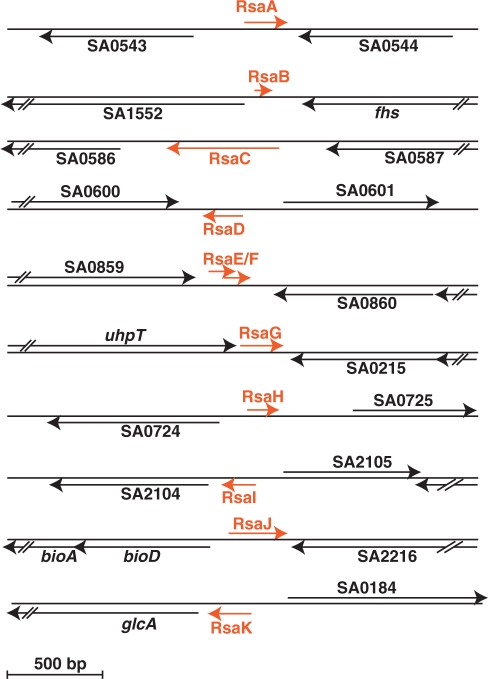
Bioinformatic analysis of the intergenic regions of Staphylococcus aureus predicted multiple regulatory regions. From this analysis, we characterized 11 novel noncoding RNAs (RsaA-K) that are expressed in several S. aureus strains under different experimental conditions. Many of them accumulate in the late-exponential phase of growth. All ncRNAs are stable and their expression is Hfq-independent. The transcription of several of them is regulated by the alternative sigma B factor (RsaA, D and F) while the expression of RsaE is agrA-dependent. Six of these ncRNAs are specific to S. aureus, four are conserved in other Staphylococci, and RsaE is also present in Bacillaceae. Transcriptomic and proteomic analysis indicated that RsaE regulates the synthesis of proteins involved in various metabolic pathways. Phylogenetic analysis combined with RNA structure probing, searches for RsaE-mRNA base pairing, and toeprinting assays indicate that a conserved and unpaired UCCC sequence motif of RsaE binds to target mRNAs and prevents the formation of the ribosomal initiation complex. This study unexpectedly shows that most of the novel ncRNAs carry the conserved C-rich motif, suggesting that they are members of a class of ncRNAs that target mRNAs by a shared mechanism.
Figures
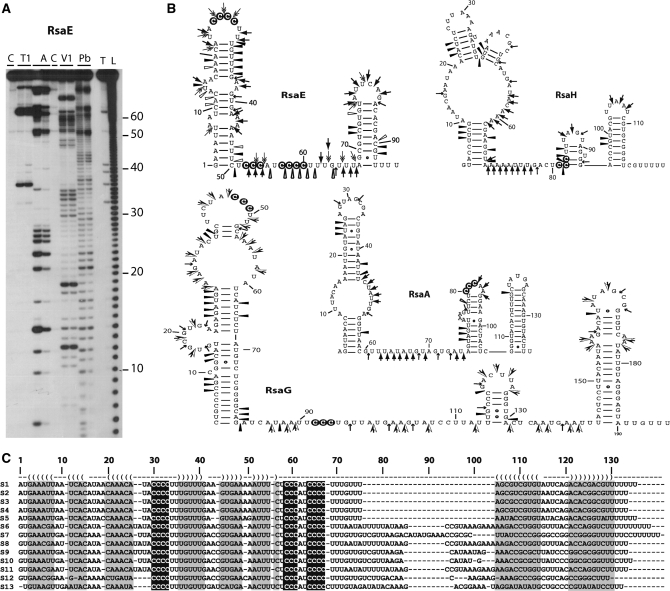
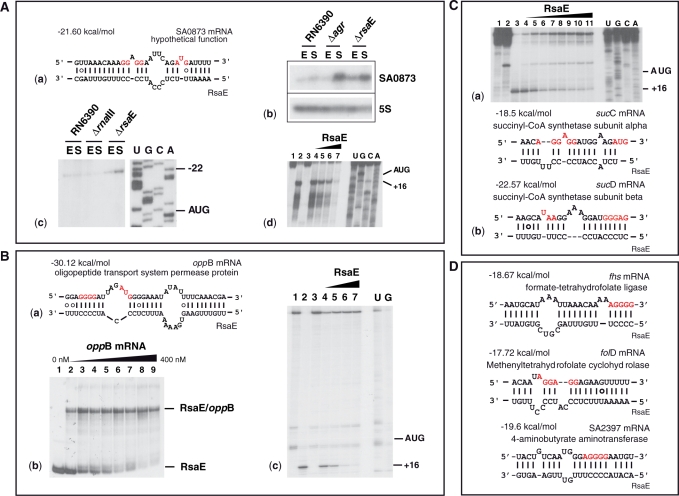
 ), RNase A (
), RNase A ( ), RNase T2 (
), RNase T2 ( ), Lead-cleavages (
), Lead-cleavages ( ) and RNase V1: (
) and RNase V1: ( ) moderate and (
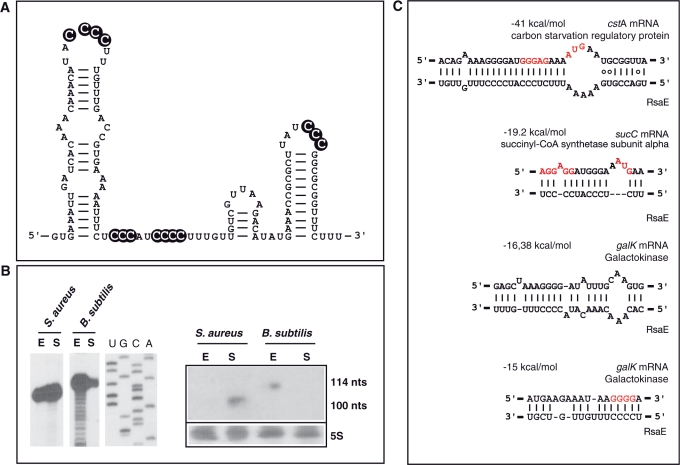
) moderate and ( ) strong cleavages. The cytosines of the C-rich motif are encircled in black. (C) Sequence and structure alignment of RsaE coming from various Staphylococci and Bacillaceae: S1, S. aureus; S2, S. epidermidis; S3, S. saprophyticus; S4, S. haemolyticus; S5, Macrococcus caseolyticus; S6, B. anthracis; S7, B. amyloliquefaciens; S8, B. cereus; S9, B. licheniformis; S10, B. subtilis; S11, B. thuringiensis; S12, Geobacillus thermodenitrificans; S13, Oceanobacillus iheyensis. The helices are shown in grey. The C-rich conserved residues are highlighted in black. The alignment was done with the PARADISE platform (
) strong cleavages. The cytosines of the C-rich motif are encircled in black. (C) Sequence and structure alignment of RsaE coming from various Staphylococci and Bacillaceae: S1, S. aureus; S2, S. epidermidis; S3, S. saprophyticus; S4, S. haemolyticus; S5, Macrococcus caseolyticus; S6, B. anthracis; S7, B. amyloliquefaciens; S8, B. cereus; S9, B. licheniformis; S10, B. subtilis; S11, B. thuringiensis; S12, Geobacillus thermodenitrificans; S13, Oceanobacillus iheyensis. The helices are shown in grey. The C-rich conserved residues are highlighted in black. The alignment was done with the PARADISE platform (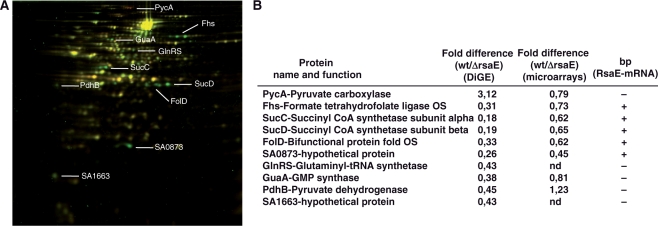

References
-
- Vogel J, Wagner EGH. Target identification of small noncoding RNAs in bacteria. Curr. Opin. Microbiol. 2007;10:262–270. - PubMed
-
- Toledo-Arana A, Repoila F, Cossart P. Small noncoding RNAs controlling pathogenesis. Curr. Opin. Microbiol. 2007;10:182–188. - PubMed
-
- Vogel J. A rough guide to the non-coding RNA world of Salmonella. Mol. Microbiol. 2009;71:1–11. - PubMed
-
- Taubes G. The bacteria fight back. Science. 2008;321:356–361. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
