

The trypanosome Rab-related proteins RabX1 and RabX2 play no role in intracellular trafficking but may be involved in fly infectivity
- PMID: 19787065
- PMCID: PMC2748683
- DOI: 10.1371/journal.pone.0007217
The trypanosome Rab-related proteins RabX1 and RabX2 play no role in intracellular trafficking but may be involved in fly infectivity
Abstract
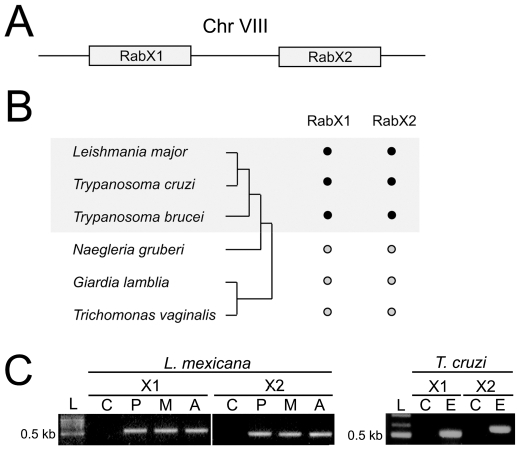
Background: Rab GTPases constitute the largest subgroup of the Ras superfamily and are primarily involved in vesicle targeting. The full extent of Rab family function is unexplored. Several divergent Rab-like proteins are known but few have been characterized. In Trypanosoma brucei there are sixteen Rab genes, but RabX1, RabX2 and RabX3 are divergent within canonical sequence regions. Where known, trypanosome Rab functions are broadly conserved when orthologous relationships may be robustly established, but specific functions for RabX1, X2 and X3 have yet to be determined. RabX1 and RabX2 originated via tandem duplication and subcellular localization places RabX1 at the endoplasmic reticulum, while RabX2 is at the Golgi complex, suggesting distinct functions. We wished to determine whether RabX1 and RabX2 are involved in vesicle transport or other cellular processes.
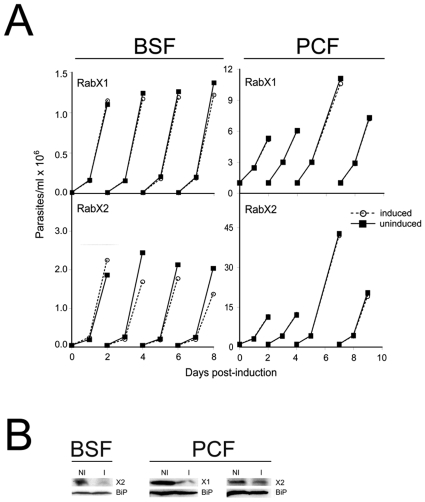
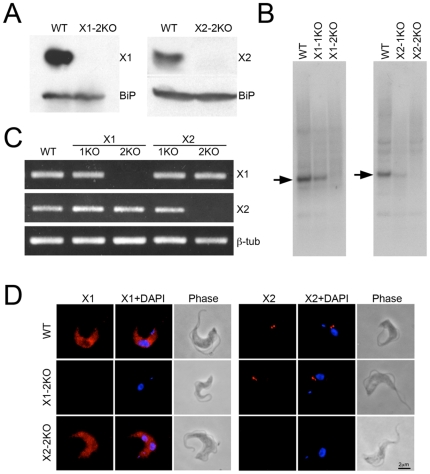
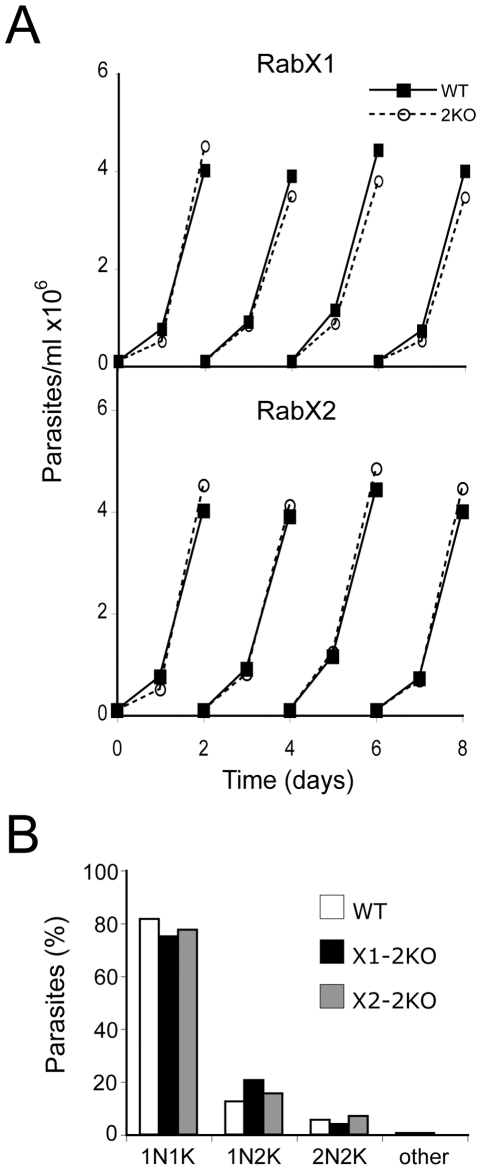
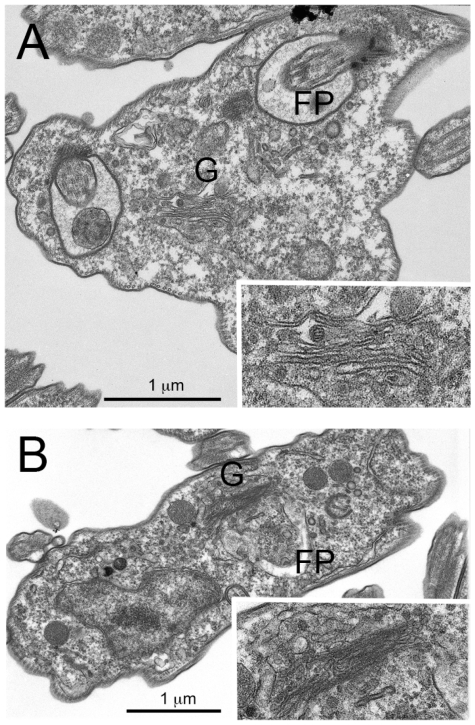
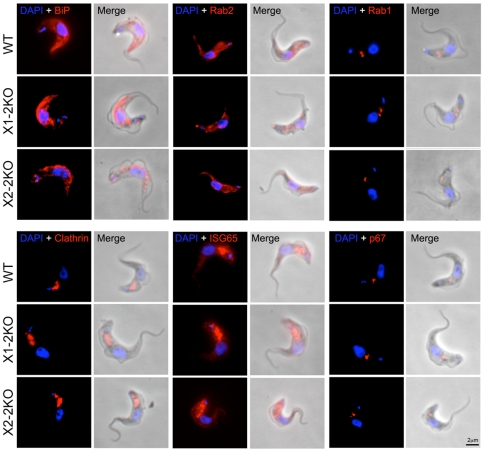
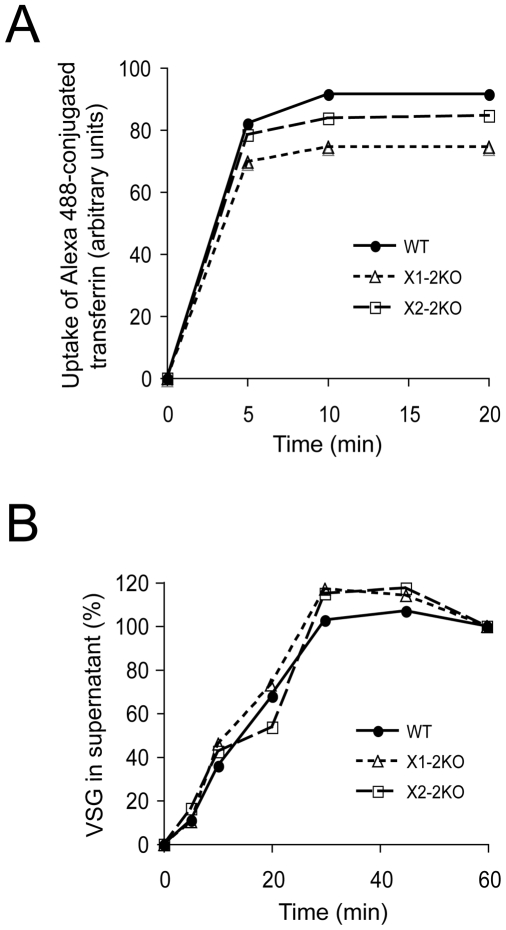
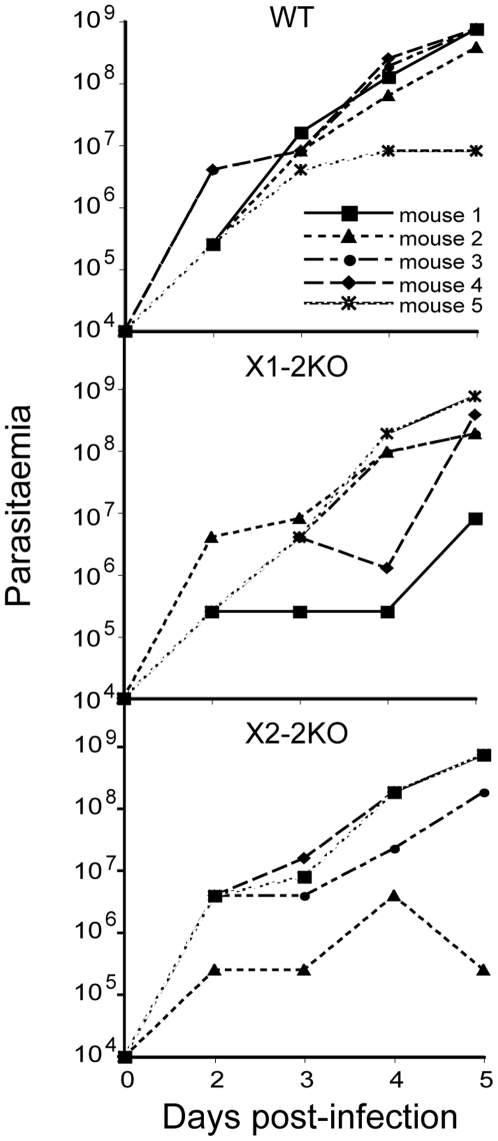
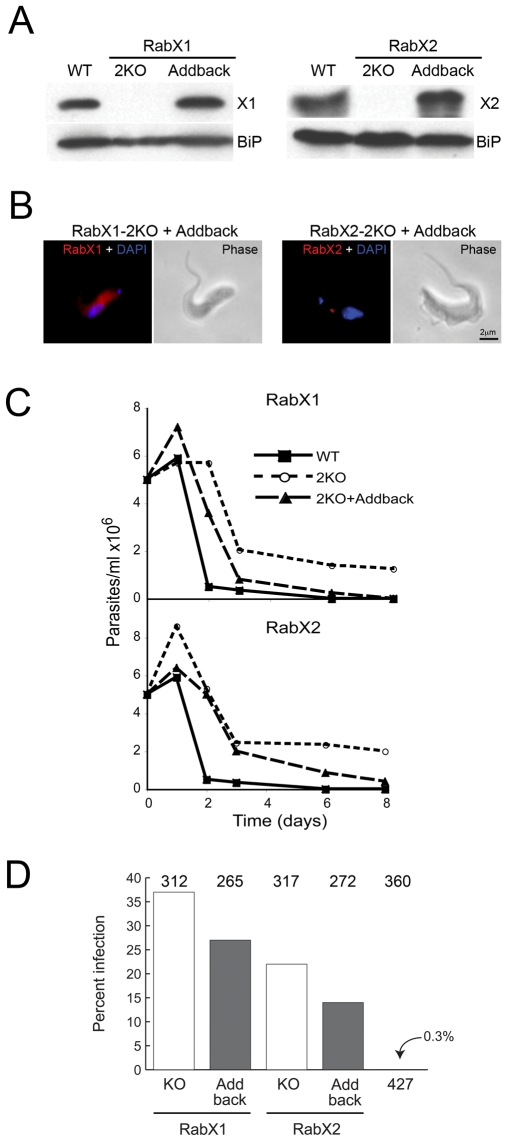
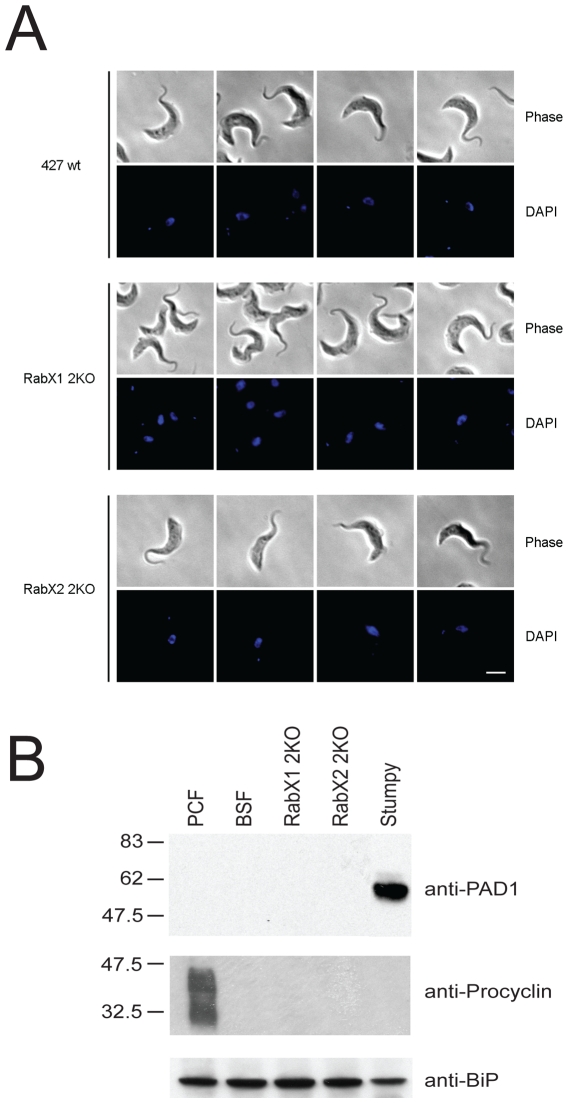
Methodology/principal findings: Using comparative genomics we find that RabX1 and RabX2 are restricted to trypanosomatids. Gene knockout indicates that RabX1 and RabX2 are non-essential. Simultaneous RNAi knockdown of both RabX1 and RabX2, while partial, was also non-lethal and may suggest non-redundant function, consistent with the distinct locations of the proteins. Analysis of the knockout cell lines unexpectedly failed to uncover a defect in exocytosis, endocytosis or in the morphology or location of multiple markers for the endomembrane system, suggesting that neither RabX1 nor RabX2 has a major role in intracellular transport. However, it was apparent that RabX1 and RabX2 knockout cells displayed somewhat enhanced survival within flies.
Conclusions/significance: RabX1 and RabX2, two members of the trypanosome Rab subfamily, were shown to have no major detectable role in intracellular transport, despite the localization of each gene product to highly specific endomembrane compartments. These data extend the functional scope of Rab proteins in trypanosomes to include non-canonical roles in differentiation-associated processes in protozoa.
Conflict of interest statement
Figures
References
-
- Barrett MP, Burchmore RJ, Stich A, Lazzari JO, Frasch AC, et al. The trypanosomiases. Lancet. 2003;362:1469–1480. - PubMed
-
- Reuner B, Vassella E, Yutzy B, Boshart M. Cell density triggers slender to stumpy differentiation of Trypanosoma brucei bloodstream forms in culture. Mol Biochem Parasitol. 1997;90:269–280. - PubMed
-
- Vassella E, Reuner B, Yutzy B, Boshart M. Differentiation of African trypanosomes is controlled by a density sensing mechanism which signals cell cycle arrest via the cAMP pathway. J Cell Sci. 1997;110:2661–2671. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
