

Bioactive and structural metabolites of pseudomonas and burkholderia species causal agents of cultivated mushrooms diseases
- PMID: 19787100
- PMCID: PMC2746572
Bioactive and structural metabolites of pseudomonas and burkholderia species causal agents of cultivated mushrooms diseases
Abstract
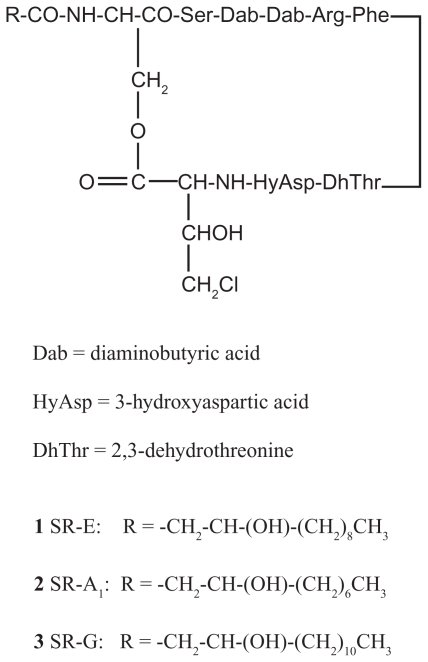
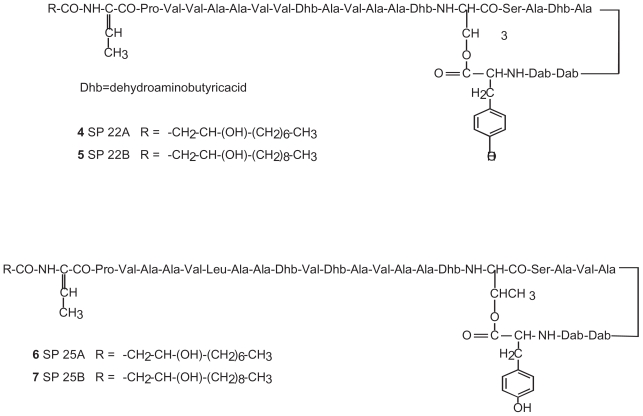
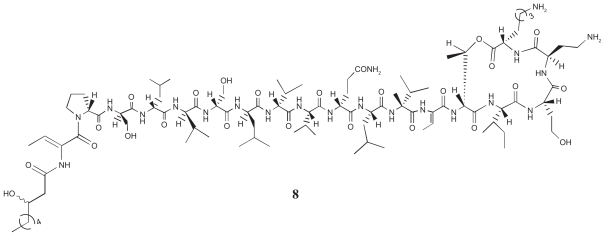
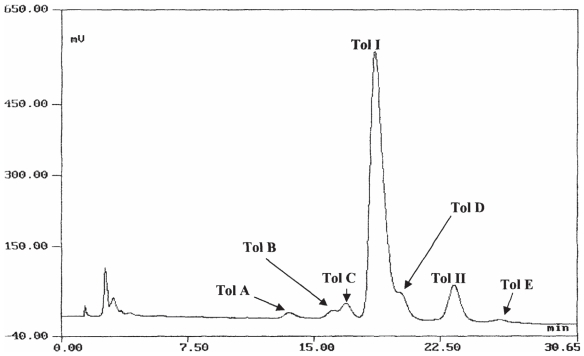
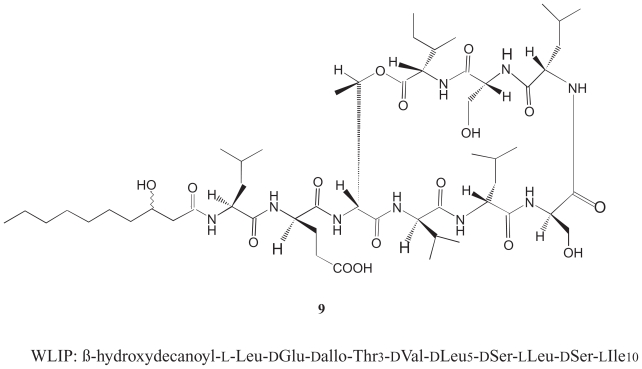
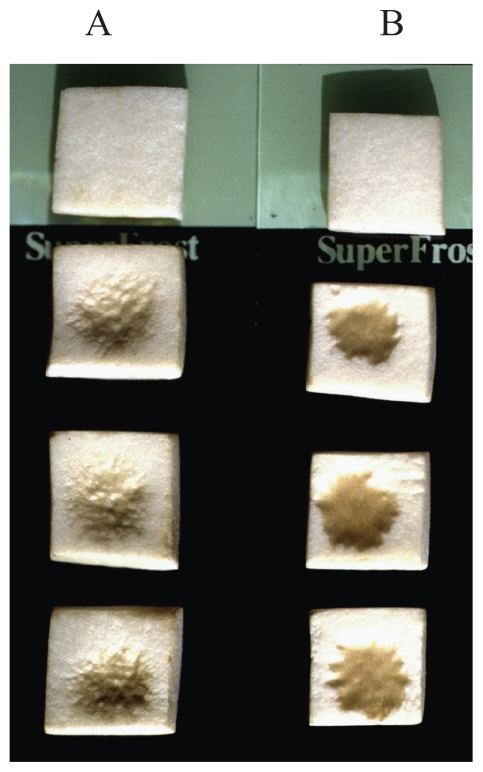
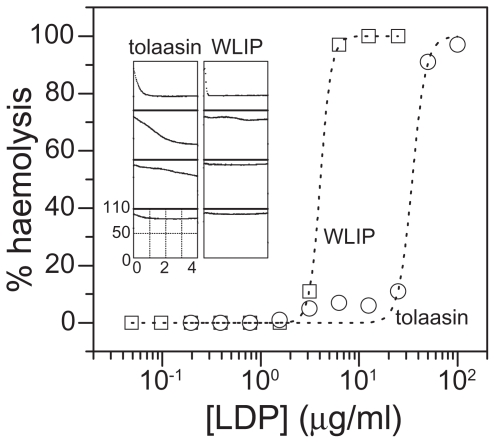
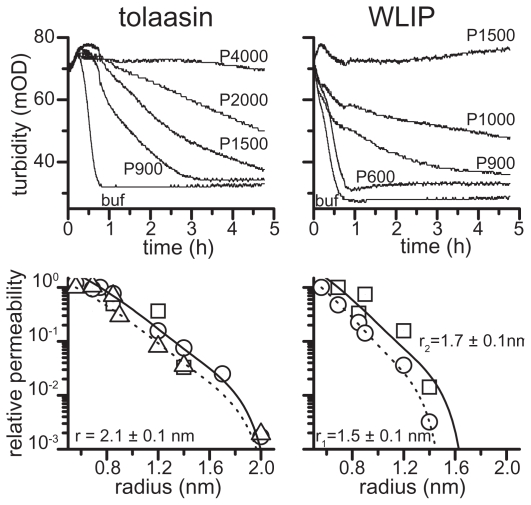
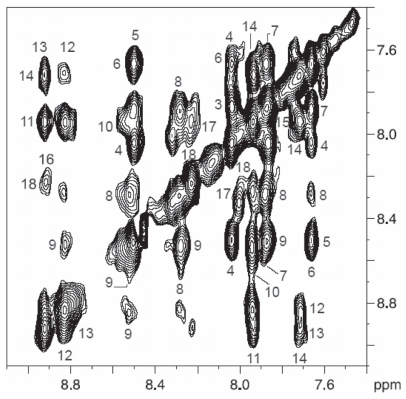
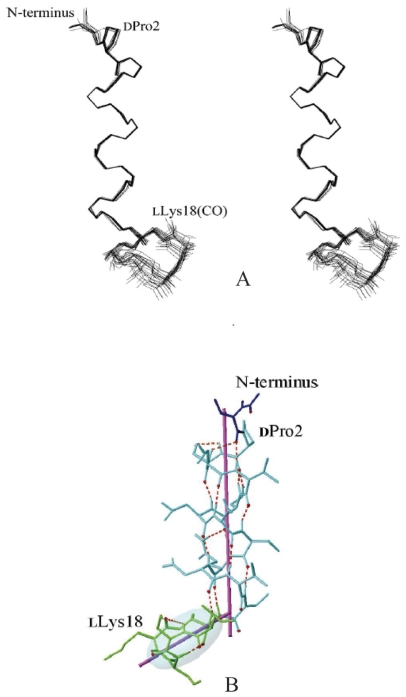
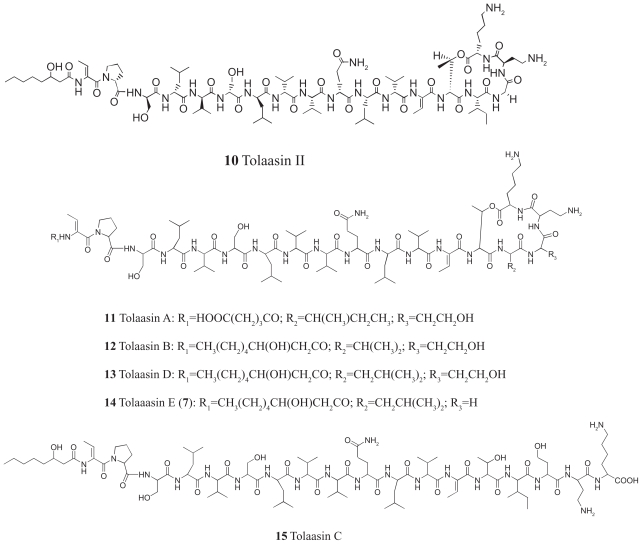
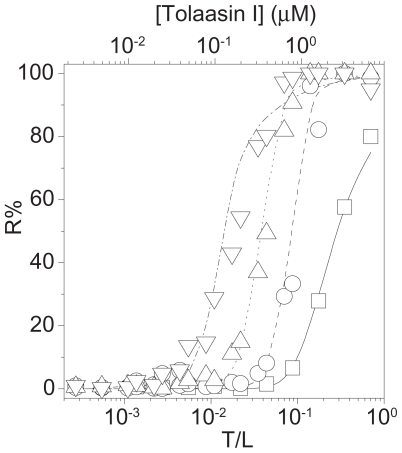
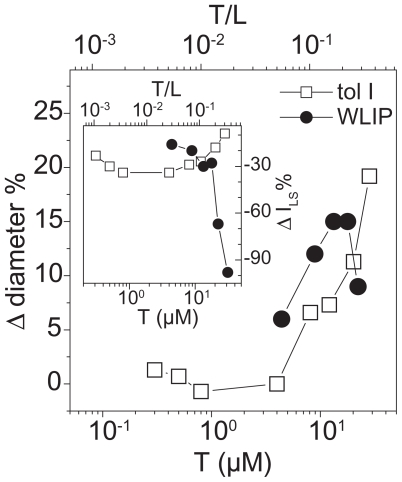
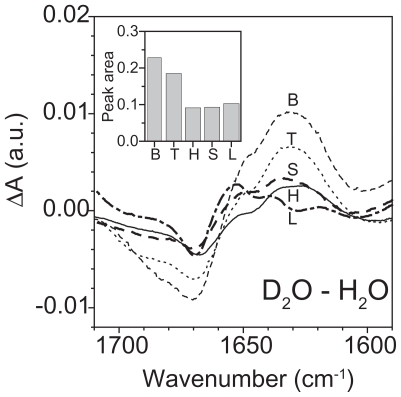
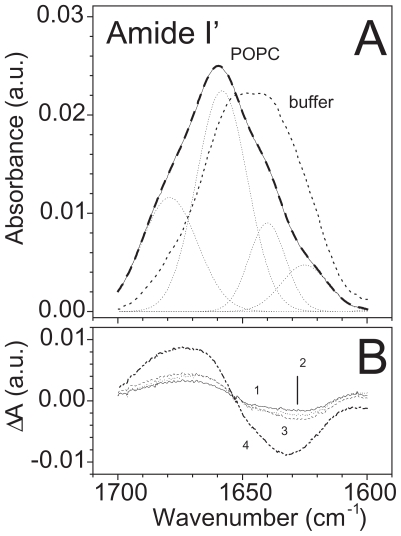
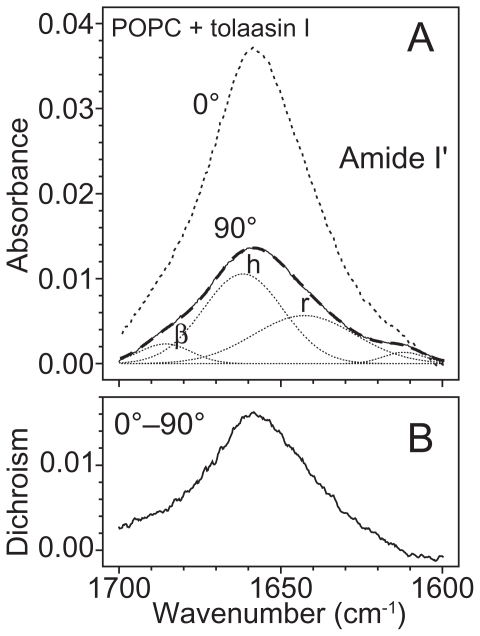
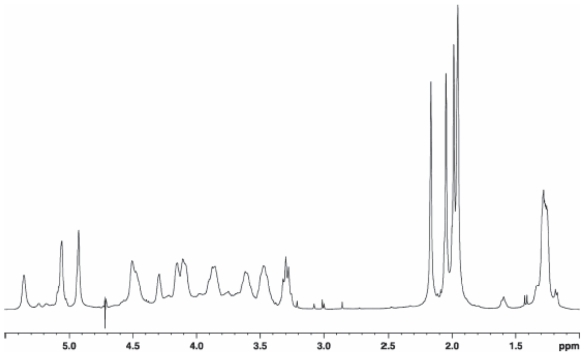
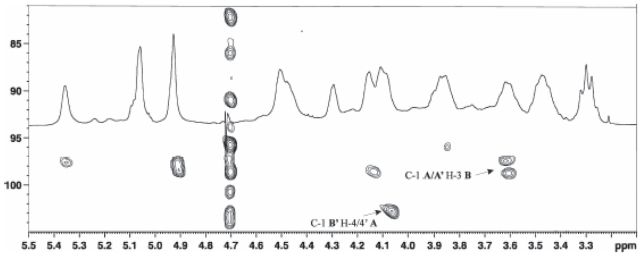
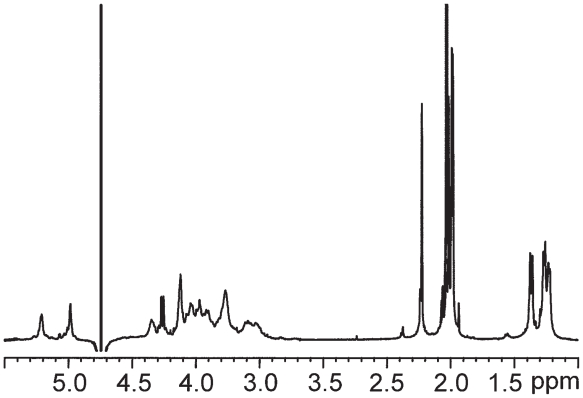
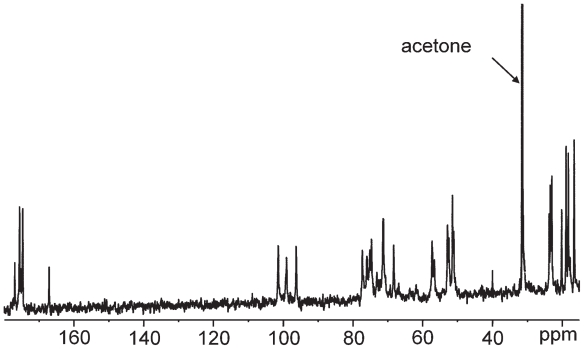
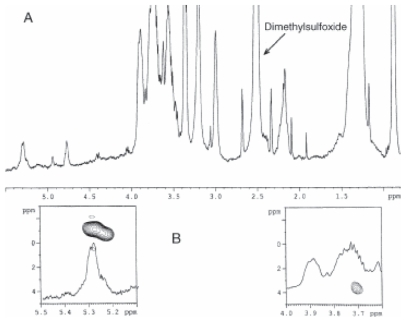
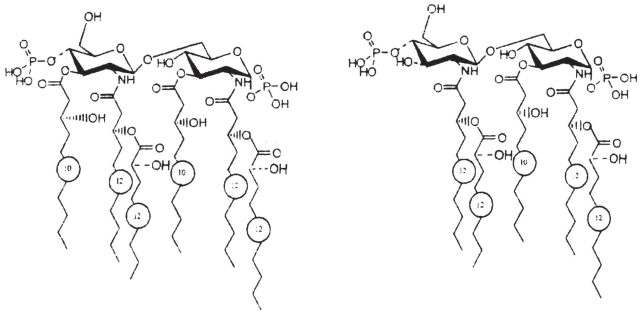
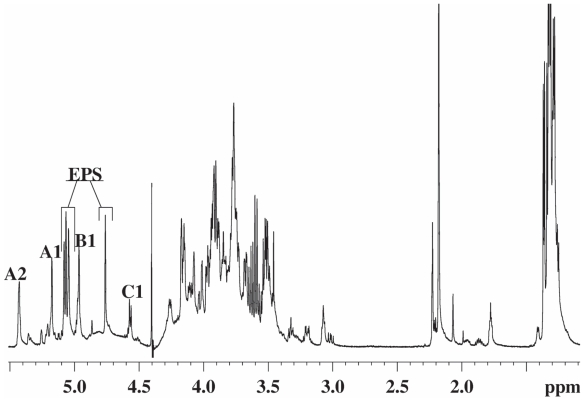
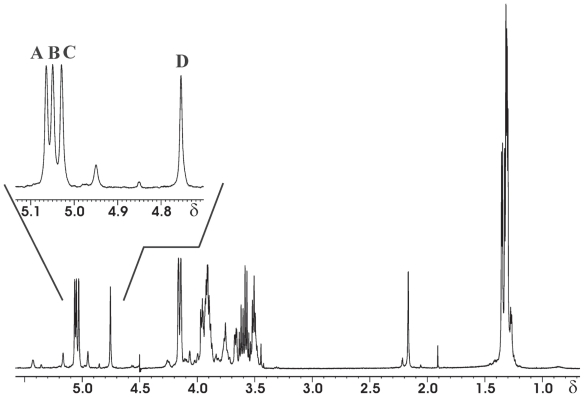
Pseudomonas tolaasii, P. reactans and Burkholderia gladioli pv. agaricicola, are responsible of diseases on some species of cultivated mushrooms. The main bioactive metabolites produced by both Pseudomonas strains are the lipodepsipeptides (LDPs) tolaasin I and II and the so called White Line Inducing Principle (WLIP), respectively, LDPs which have been extensively studied for their role in the disease process and for their biological properties. In particular, their antimicrobial activity and the alteration of biological and model membranes (red blood cell and liposomes) was established. In the case of tolaasin I interaction with membranes was also related to the tridimensional structure in solution as determined by NMR combined with molecular dynamic calculation techniques. Recently, five news minor tolaasins, tolaasins A-E, were isolated from the culture filtrates of P. tolaasii and their chemical structure was determined by extensive use of NMR and MS spectroscopy. Furthermore, their antimicrobial activity was evaluated on target micro-organisms (fungi-including the cultivated mushrooms Agaricus bisporus, Lentinus edodes, and Pleurotus spp.-chromista, yeast and bacteria). The Gram positive bacteria resulted the most sensible and a significant structure-activity relationships was apparent. The isolation and structure determination of bioactive metabolites produced by B. gladioli pv. agaricicola are still in progress but preliminary results indicate their peptide nature. Furthermore, the exopolysaccharide (EPS) from the culture filtrates of B. gladioli pv. agaricicola, as well as the O-chain and lipid A, from the lipopolysaccharide (LPS) of the three bacteria, were isolated and the structures determined.
Keywords: P. reactans and Burkholderia gladioli pv. agaricicola; Pseudomonas tolaasii; antimicrobial activity; cultivated mushrooms and bacterial diseases; exopolysaccharides and lipopolysaccharides; lipodepsipetides; mycopathogenic bacteria; permeabilising effects on membranes.
Figures
References
-
- Andreu D, Rivas L. Animal antimicrobial peptides: an overview. Biopolymers. 1998;47:415–3. - PubMed
-
- Arrebola E, Cazorla FM, Duran VE, et al. Mangotoxin: a novel antimetabolite toxin produced by Pseudomonas syringae inhibiting ornithine/arginine biosynthesis. Physiol. Mol. Plant Pathol. 2003;63:117–27.
-
- Atkey PT, Fermor TR, Lincoln SP. In: Maher MJ, editor. Proceedings of the 13th International Congress on The Science and Cultivation of Edible Fungi; Rotterdam: A. A. Balkema; 1991. p. 431.
-
- Avrahami D, Shai Y. Conjugation of a magainin analogue with lipophilic acids controls hydrophobicity, solution assembly, and cell selectivity. Biochemistry. 2002;41:2254–63. - PubMed
-
- Ballio A, Bossa F, Di Giorgio A, et al. Solution conformation of the Pseudomonas syringae pv. syringae phytotoxic lipodepsipeptide syringopeptin 25-A. Two-dimensional NMR, distance geometry and molecular dynamics. Eur. J. Biochem. 1995;234:747–58. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous