

Hematopoietic progenitor kinase 1 is a critical component of prostaglandin E2-mediated suppression of the anti-tumor immune response
- PMID: 19787351
- PMCID: PMC2798028
- DOI: 10.1007/s00262-009-0761-0
Hematopoietic progenitor kinase 1 is a critical component of prostaglandin E2-mediated suppression of the anti-tumor immune response
Abstract
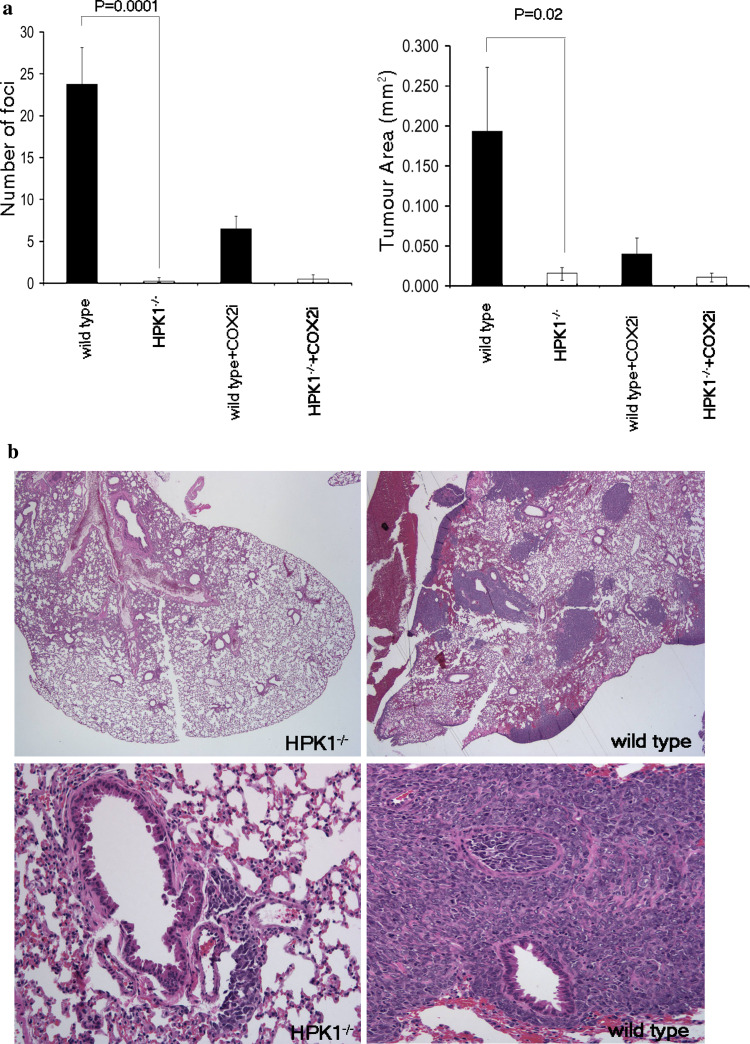
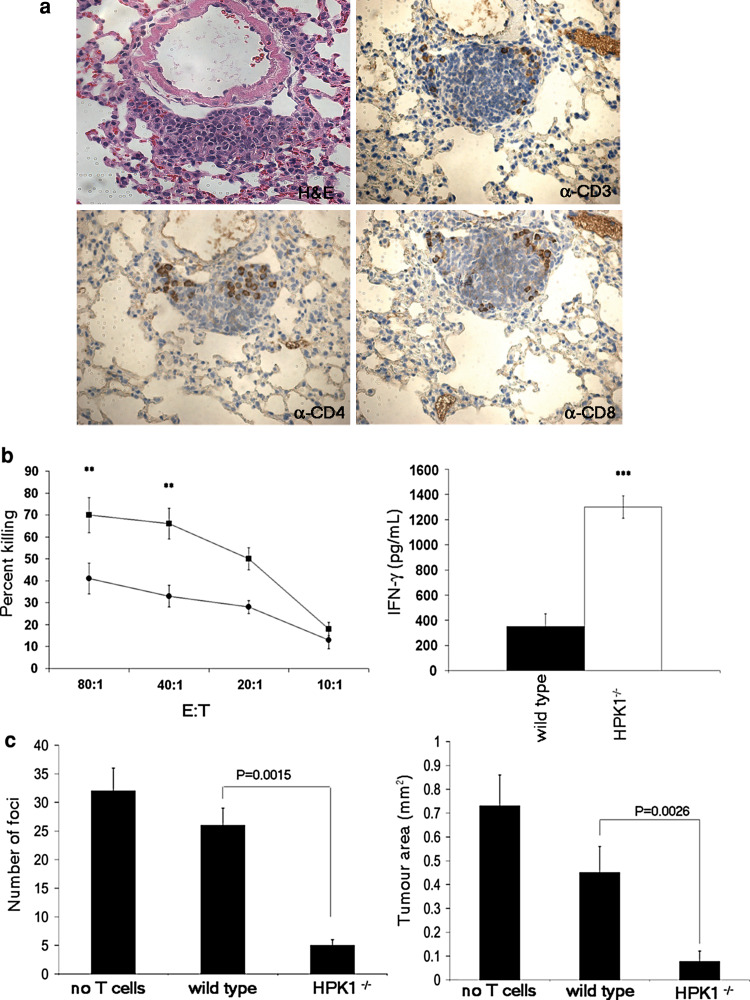
Lung cancer is the leading cause of cancer-related mortality in the world, resulting in over a million deaths each year. Non-small cell lung cancers (NSCLCs) are characterized by a poor immunogenic response, which may be the result of immunosuppressive factors such as prostaglandin E2 (PGE(2)) present in the tumor environment. The effect of PGE(2) in the suppression of anti-tumor immunity and its promotion of tumor survival has been established for over three decades, but with limited mechanistic understanding. We have previously reported that PGE(2) activates hematopoietic progenitor kinase 1 (HPK1), a hematopoietic-specific kinase known to negatively regulate T-cell receptor signaling. Here, we report that mice genetically lacking HPK1 resist the growth of PGE(2)-producing Lewis lung carcinoma (LLC). The presence of tumor-infiltrating lymphocytes (TILs) and T-cell transfer into T cell-deficient mice revealed that tumor rejection is T cell mediated. Further analysis demonstrated that this may be significantly due to the ability of HPK1 (-/-) T cells to withstand PGE(2)-mediated suppression of T-cell proliferation, IL-2 production, and apoptosis. We conclude that PGE(2) utilizes HPK1 to suppress T cell-mediated anti-tumor responses.
Figures



References
-
- Arnold R, Patzak IM, Neuhaus B, Vancauwenbergh S, Veillette A, Van Lint J, Kiefer F. Activation of hematopoietic progenitor kinase 1 involves relocation, autophosphorylation, and transphosphorylation by protein kinase D1. Mol Cell Biol. 2005;25:2364–2383. doi: 10.1128/MCB.25.6.2364-2383.2005. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
