

Review
doi: 10.1021/cr900077w.
Coordination chemistry of bacterial metal transport and sensing
Affiliations
- PMID: 19788177
- PMCID: PMC2783614
- DOI: 10.1021/cr900077w
Item in Clipboard
Review
Coordination chemistry of bacterial metal transport and sensing
Chem Rev.
2009 Oct.
No abstract available
Figures
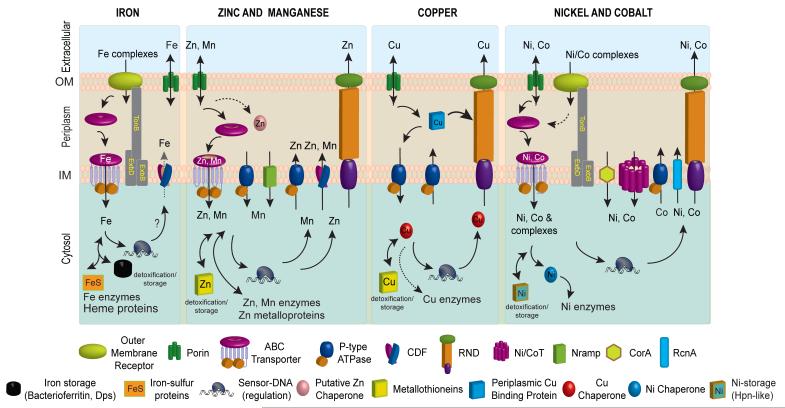
Schematic metal homeostasis models for iron, zinc and manganese, copper, nickel and cobalt, shown specifically in gram-negative bacteria. Homeostasis of molybdate and tungstate oxyanions are not shown, due primarily to a lack of knowledge of these systems, outside of uptake (Section 2.5) and cytosolic sensing (Section 3.8.1). This schematic is not representative for any one bacterium nor is it meant to be exhaustive, but is instead simply designed to convey the potential fates of individual metal ions in distinct cellular compartments. Not all bacteria have all components of each homeostasis system indicated. The double-headed arrows are meant to illustrate that metals can move and in and out of target protein destinations in response to proteome remodeling. Specific protein designations for individual homeostasis components are indicated in Figures 2 and 3. A putative chaperone shown for Zn(II) is YodA/ZinT, while actual metallochaperones for Cu(I) (Atx1 and CopZ407), and for Ni(II) metalloenzymes urease (UreE10) or Ni-Fe hydrogenases (e.g., HypA10) are also shown. Iron metallochaperones for Fe-S cluster assembly are not shown for clarity. Fe(II) efflux through YiiP has not yet firmly established biochemically. The cytosolic Cu(I) quota for a non-photosynthetic bacterium may well be vanishingly small; as a result, transfer from a Cu(I) chaperone is indicated by the dashed double-headed arrow.
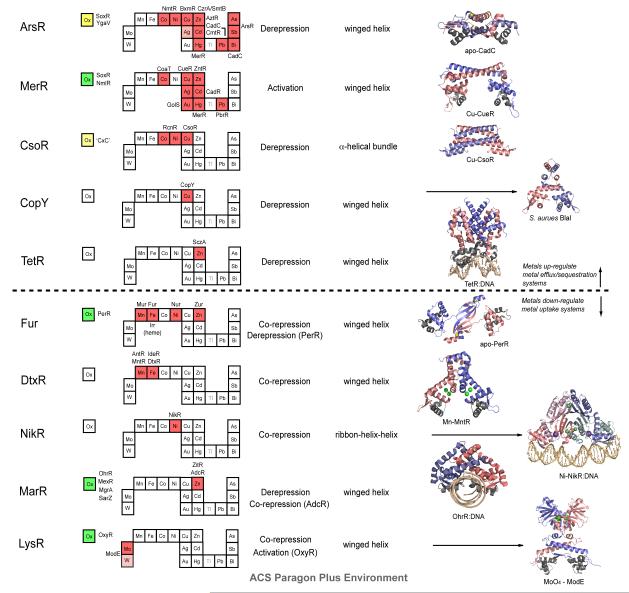
Structural families of metalloregulatory proteins. For each family, boxes for metals that are known to be sensed are shaded red on the abbreviated periodic table, while green boxes on the left denote known family members that are known to sense cytosolic oxidative stress. Boxes identifying putative metal sensors and non-metal sensing oxidative stress regulators are shaded pink or yellow, respectively. The four-letter designations for individual proteins that perform the function listed in the nearby box are given (see text for details). The mechanism of regulation of gene expression is indicated as is the DNA-binding domain that mediates operator-promoter DNA binding. Ribbon representations of selected representative members are shown on the right with individual protomers shaded red and blue in each case. Structures are from top to bottom: 1) apo S. aureus pI258 CadC with structural α5-Zn(II) ions shaded yellow (1U2W pdb code); 2) E. coli Cu(I)-sensor CueR with regulatory Cu(I) ions in red (1Q05); 3) M. tuberculosis Cu(I)-sensor CsoR with regulatory Cu(I) ions shaded red (2HH7); 4) S. aureus BlaI as a model for Enterococcus CopY (1SD4); 5) TetR-Tc-Mg DNA complex structure as model for S. pneumonia SczA (3CDL); 6) apo B. subtilis PerR with structural Zn(II) ions in yellow (2FE3); 7) B. subtilis MntR with MnA/MnC binuclear cluster ions in green (2F5F); 8) Ni(II)-bound E. coli NikR-nik operator DNA complex with high affinity Ni(II) ions shown in green and regulatory K+ ions in purple (2HZV); 9) B. subtilis OhrR-DNA complex (1Z9C); 10) Molybdate sensor ModE with molybdate shaded green (1O71). Adapted with permission from Reference . Copyright 2007 Royal Society of Chemistry.
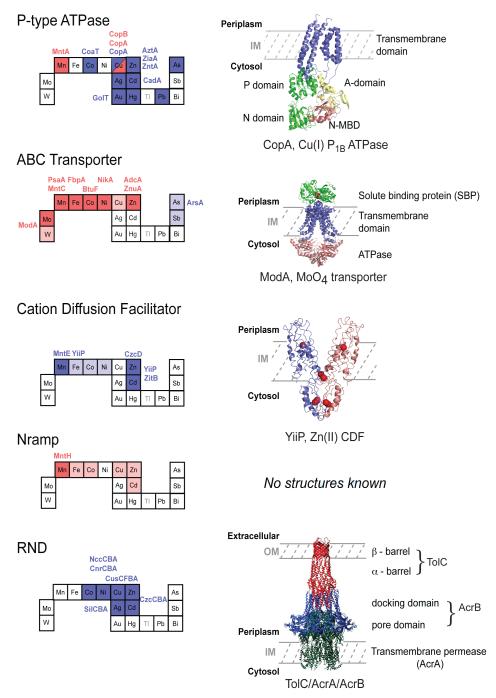
Structural families of metal transporters. Individual elements on the abbreviated periodic table are shaded red if the are specifically imported by a member of that particular family or blue if they are effluxed. Metal ions that are transported non-specifically are shaded pink for uptake or light blue for efflux. The four-letter designations for individual proteins that perform the indicated function are given; this list is not meant to be comprehensive but rather shows representative members that have been structurally and/or functionally characterized (see text for details). Ribbon representations of crystallographic structures of one member of each family summarizes salient structural features of each transporter. The structures shown are 1) Archaeoglobus fulgidus CopA monomer, a Cu(I)-selective P1B-type ATPase (2VOY); 2) Archaeoglobus fulgidus MoO4−—ABC transporter with MoO4− in red (2ONK); 3) E. coli Zn(II) transporting CDF proteins YiiP with zinc atoms shaded red (2QFI); 4) There are no reported structures for an Nramp transporter; 5) Combined hypothetical structure of an RND complex. This panel adapted with permission from Reference . Copyright 2004 Elsevier Limited.
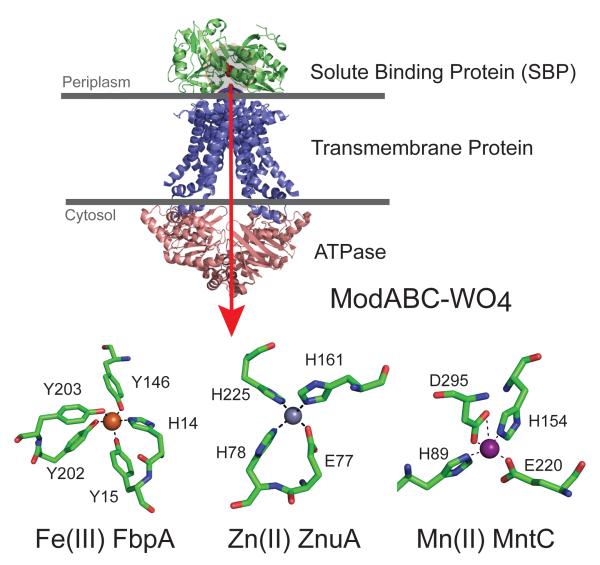
Structure of a representative ABC importer, E. coli ModABC with the direction of transport shown by the red arrow. The green subunit ModA is the solute binding protein (SBP), with the position of the WO42− anion shaded red and highlighted in the gray circle. Other SBPs adopt a global structure similar to ModA but feature distinct metal coordination sites as shown for Fe(III)-, Zn(II)- and Mn(II)-specific SBPs (lower part of figure). Coordination sites are shown for the Fe(III)-specific SBP Campylobacter jejuni FbpA (1Y4T), a Zn(II) -SBP E. coli ZnuA (2OSV), the Mn(II)-SBP Synechocystis 6803 MntC (1XVL).
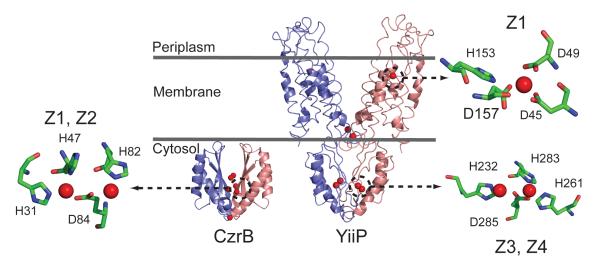
Structural comparison of the intact Zn(II) CDF transporter E. coli YiiP (2QFI) with the cytosolic domain of the homolog T. thermophilus CzrB (3BYR). Each structure highlights the Zn(II) coordination environments for zinc in both YiiP and CzrB. The structures of CzrB and YiiP were refined with 3 and 4 Zn(II) ions per protomer, respectively. The cytosolic protein CzrB contains a divalent metal center (Z1 and Z2) that is roughly, but not precisely analogous, to the Z3 and Z4 sites in YiiP. The transmembrane Z1 site in E. coli YiiP is also shown with a key ligand D157 highlighted.
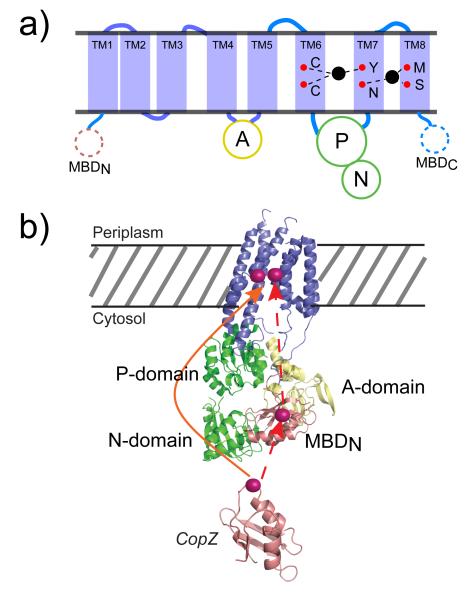
(a) Schematic representation of the topology of a heavy metal P1B-type ATPase transporter. TM, transmembrane helix; MBD, metal-binding domain; A, actuator domain; P, phosphorylation domain; N, nucleotide binding domain; red spheres, approximate positions of mapped residues important for Cu(I) transport by A. fulgidus CopA; black spheres, metal ion bound to the transmembrane binding site(s) in a manner consistent with x-ray absorption studies. (b) Models for Cu(I) delivery to the transmembrane metal binding sites in the CopA P-type ATPase from A. fulgidus. A ribbon diagram of a CopA monomer modeled on the cryo-EM structure of the CopA dimer. Transmembrane helices shown in slate, P/N domains in green, and the A domain in colored yellow. The N-terminal MBD is shown in salmon. The Cu(I)-chaperone CopZ ribbon diagram in shaded salmon is based on the structure of Enterococcus hirae CopZ (PDB ID code 1CPZ). The dashed red line symbolizes the intermediate transfer of Cu(I) from CopZ to the N-MBD, to the transmembrane metal binding site(s). The red solid line represents the direct delivery of Cu(I) in CopZ to the transmembrane metal binding site of CopA. The locations of the bond Cu(I) ions are for schematic purposes only. Adapted with permission from Reference . Copyright 2008 National Academy of Sciences U.S.A.
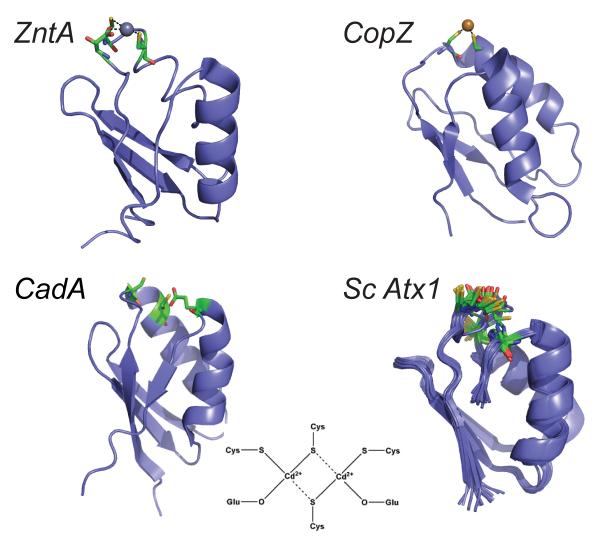
Ribbon diagrams of the N-terminal MBDs of the P-type ATPases E. coli ZntA and Listeria monocytogenes CadA compared to representative bacterial Cu(I) metallochaperones from the N-terminal domain of CopZ from B. subtilis and ScAzx1 from Synechocystis PCC 6803 (an NMR bundle). A schematic rendering of the dimeric, binuclear Cd(II) complex adopted by CadA is also shown.
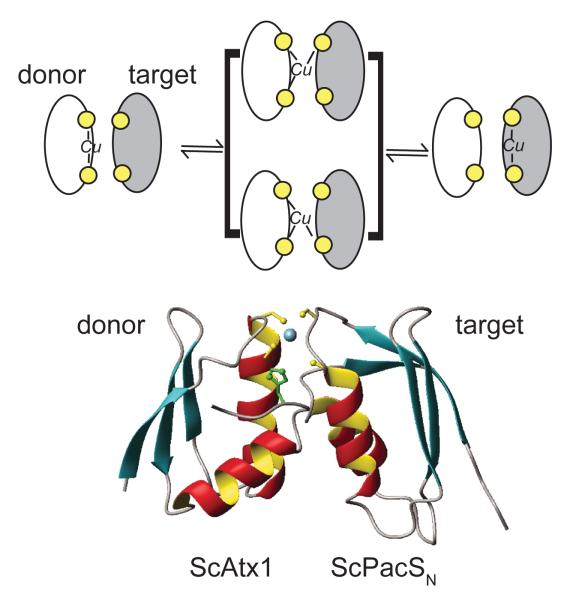
(a) Schematic model of Cu(I) exchange between a Cu(I) donor metallochaperone and a Cu(I) target protein, e.g., the MBD of a P-type ATPases. The structural intermediate shown in brackets is a transiently formed Cu(I)-cross linked intermolecular complex, a three-dimensional NMR-based model of which is shown in (b) for the complex between the Cu(I) chaperone ScAtx1 and the N-terminal MBD of PacS from Synechocystis PCC 6803. Adapted with permission from Reference . Copyright 2006 National Academy of Sciences U. S. A.
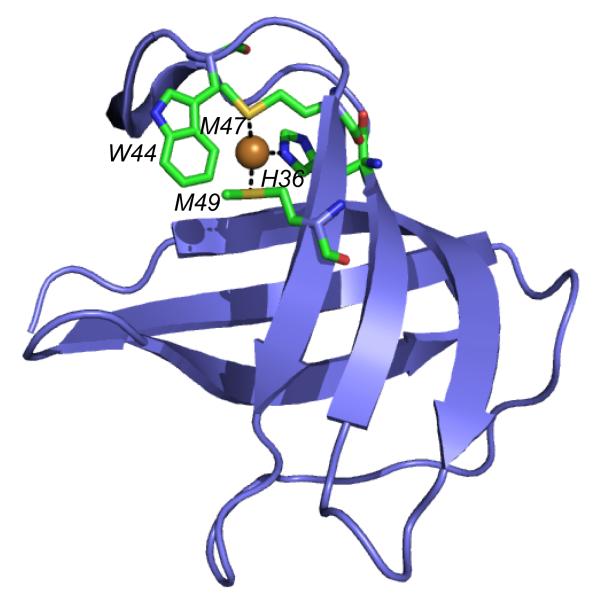
A ribbon representation of E. coli CusF highlighting the tetragonal distortion of the Met-His-Met trigonal Cu(I) plane by the indole ring of W44 which forms a classical cation-π interaction with the Cu(I) ion.,
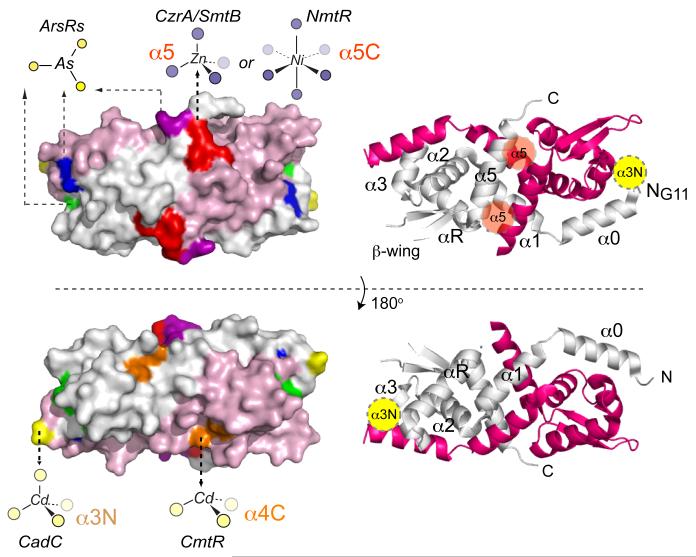
Summary of the known metal binding sites of ArsR/SmtB family repressors on the structure of S. aureus pI258 CadC homodimer. Left, spacefilling models of two views of CadC adapted from ref. with ribbon representations of the same view shown on the right. One protomer is shaded pink and the other grey, with the α-helices labeled consecutively from the N-terminus α0-α5 of the ribbon diagrams (which correspond to α1-α6 in the Ye et al. structure), along with schematic locations of the α3N (yellow) and α5 (shaded red) sensing sites on each view of the dimer. The approximate locations and schematic renderings of representative coordination complexes of distinct sensing sites are shown on the left and correspond to S. aureus pI258 CadC (yellow, α3N), E. coli plasmid R773 ArsR (green, α3), S. aureus CzrA/Synechococcus SmtB and M. tuberculosis NmtR (red, α5 and α5C, respectively), M. tuberculosis CmtR (orange, α4C), C. glutamicum ArsR1 (blue) and A. ferrooxidans ArsR (purple). The α5-3 metal site characterized in the Ni/Co sensor M. tuberculosis KmtR is not explicitly shown, but partially overlaps the α5 site. See text for details and Figure 11 for a multiple sequence alignment that highlights these metal sensor sites in the ArsR/SmtB family.
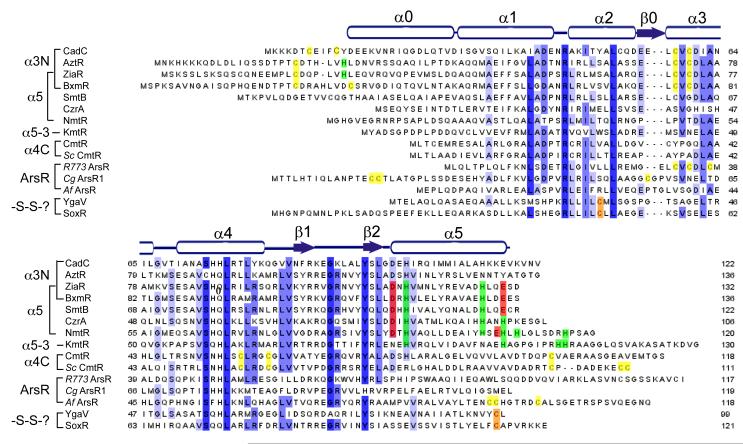
Multiple sequence alignment of ArsR/SmtB family repressors discussed here with the secondary structural units of apo-CadC shown. These secondary structural units align well with those known for S. aureus CzrA and Synechococcus SmtB. The residues known to coordinate regulatory metal ions in each sequence are shaded yellow (Cys), green (His) or red (Asp/Glu) in each sequence, with degree of residue-specific conservation at each position in the alignment indicated by the blue shading. See text for details.
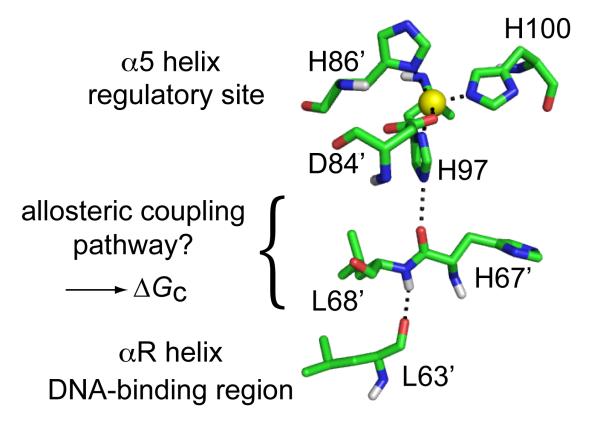
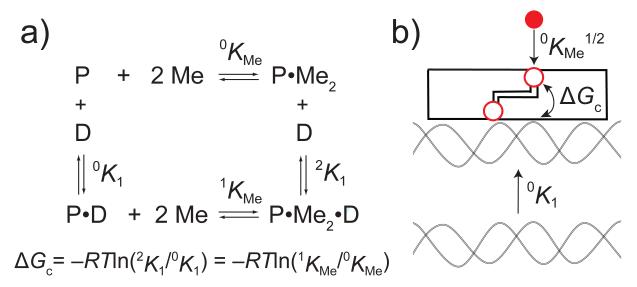
Proposed hydrogen bonding network in Zn(II)-bound CzrA that links the α5 regulatory sites to that of the DNA binding helices, which is thought to contribute directly to the magnitude of ΔGc (see Scheme 1).,
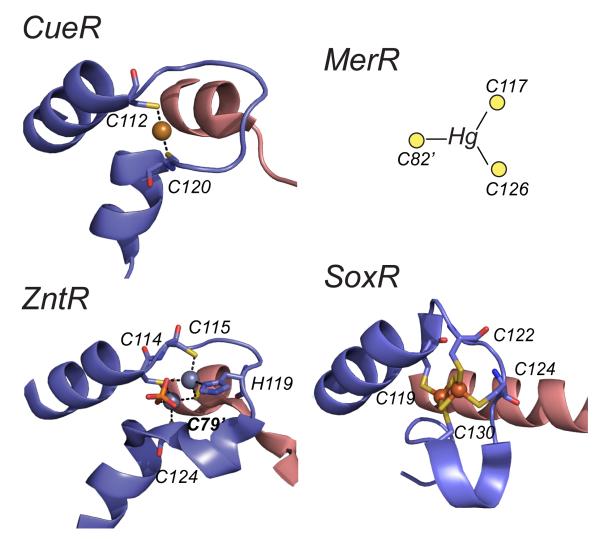
Ribbon representations of the metal binding loops of various MerR family metalloregulators. In all cases, only one of the two symmetry-related metal sites are shown with one protomer is shaded blue and the other red; annotated metal donor ligands shown in stick. The structures shown are the Cu(I) bound form of E. coli CueR, the Zn(II)2 sulfate anion (shown in red/orange)-bridged binuclear structure of E. coli ZntR, and the [2Fe-2S]2+ center of E. coli SoxR. A schematic of the single subunit-bridging Hg(II) site of Tn501 MerR consistent with spectroscopic and functional data, but of unknown structure is also shown for comparison.
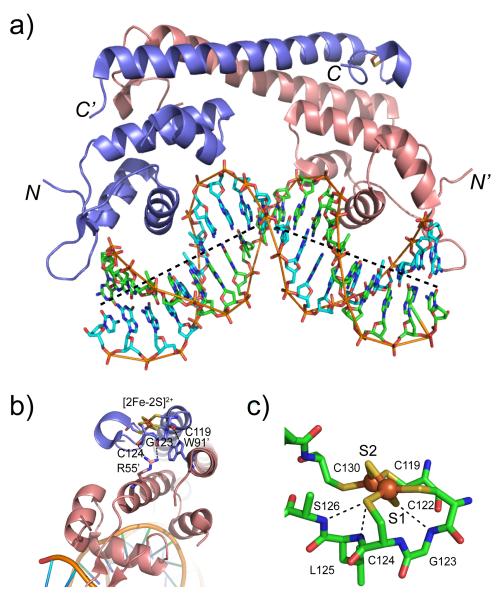
Crystallographic structure of the oxidized (activated) E. coli SoxR-DNA complex. (a) Overall view of the structure of SoxR-DNA complex showing a significant bend (~65°) in the sox operator DNA. The DNA strands are colored green and cyan and shown in stick representation; the two protomers of the SoxR homodimer are shown as ribbon structures, and shaded as in Figure 15. (b) Intersubunit hydrogen bonding interactions that link main chain carbonyl oxygen atoms from G123 and C124 in the metal binding loop that coordinates the [2Fe-2S]2+ cluster with the side chain of R55′ from the DNA binding domain of the opposite protomer. A main chain-side chain hydrogen bond between C119 and W91′ from the dimerization helix of the opposite protomer is also shown. (c) A close-up view of the [2Fe-2S]2+ cluster revealing how electrostatic interactions around the bridging S2− anion S1 may facilitate the conformational change upon reversible reduction/oxidation of the cluster.
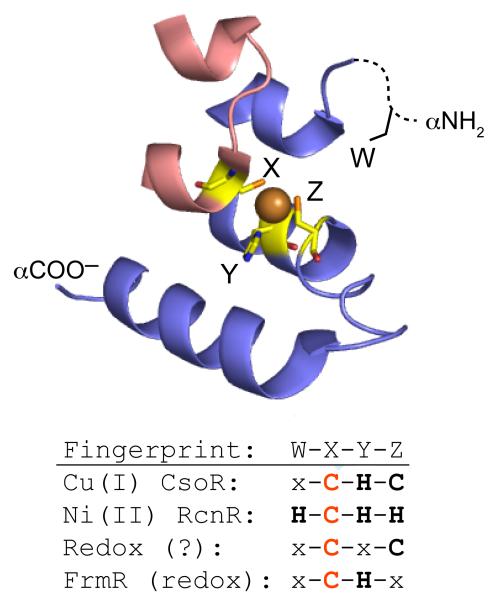
A schematic representation of the W-X-Y-Z “fingerprint” of individual CsoR/RcnR family repressors adapted from ref. . The X-Y-Z region of the fingerprint is defined by the ligands to the Cu(I) ion in Cu(I)-sensing CsoRs, corresponding to C36, H61′ and C65′ in opposite protomers of M. tuberculosis CsoR (shaded red and blue, respectively). W corresponds to H3 in E. coli RcnR which must occupy the third position relative to the M1 αNH2 group. The invariant Cys in the X position is shaded red.
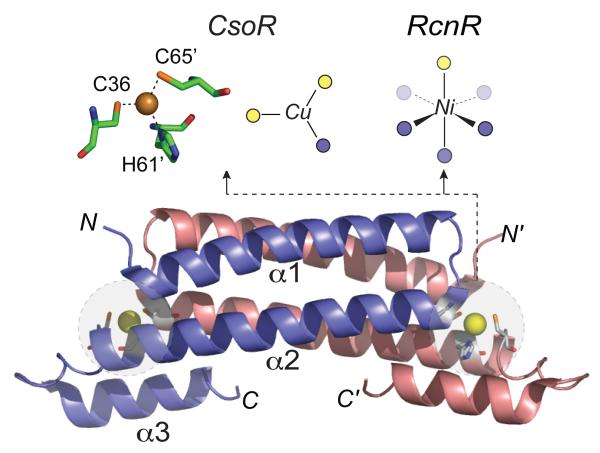
Ribbon representation of the 2.6 Å crystallographic structure of the Cu(I)-bound M. tuberculosis CsoR homodimer. The crystallographically defined structure of the Cu(I) coordination complex is shown in stick representation, while schematic representations of Cu(I) and Ni(II) complexed of CsoR and E. coli RcnR, respectively, are also shown. Cys sulfur ligands are shaded yellow while N/O ligands are given by the blue spheres. The α-helices of the blue protomer in CsoR are labeled α1-α3.
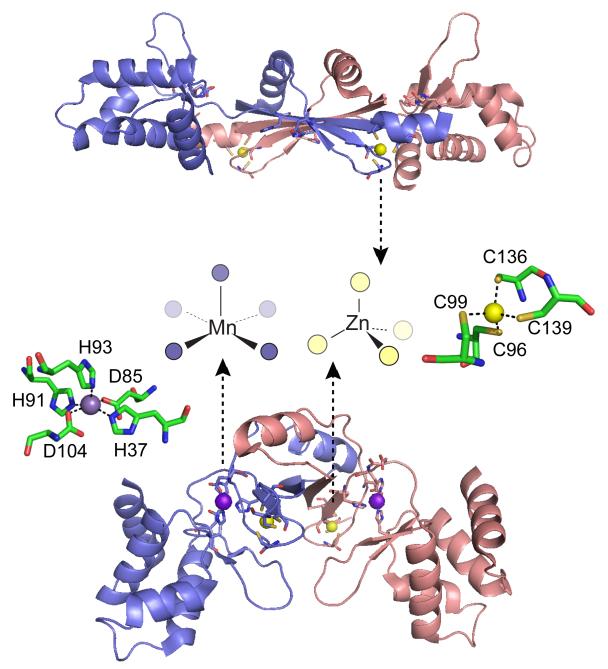
Ribbon diagrams of two crystallographic structures of B. subtilis PerR with the subunits shaded blue and red. Top, oxidized form of PerR, designated PerR-Zn-ox, in which the regulatory metal sites are empty and H37 and H91 are modeled as 2-oxo-histidine residues. Each of two structural Zn(II) ions are bound to the homodimer in a tetrathiolate, tetrahedral coordination complex that is conserved in some but not all Fur family members. Bottom, Mn(II)-activated PerR, denoted PerR-Zn-Mn, in which the H2O2 sensing or regulatory site is formed by a square pyramidally coordinated Mn(II) or Fe(II) atom by H37 from the winged helical DNA binding domain (on the periphery of the homodimer), D85 from the connecting linker, and H91, H93 and D104 from the dimerization domain (middle), all from the same protomer. The symmetry-related metal ligands are also shown on the opposite subunit. H37 is oxidized to 2-oxo-His in PerR-Zn-oxo (shown), as is H91., The structural model of PerR-Zn-ox (top) superimposes on apo-PerR-Zn.
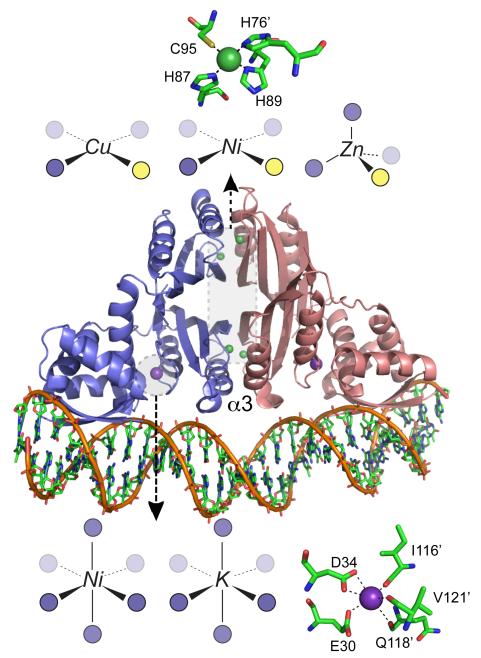
Ribbon representation of the crystallographic structure of the E. coli NikR-DNA complex with Ni(II) (shaded green) bound at the high affinity C-terminal sites and K+ ions (shaded purple) bound at the low affinity sites in close proximity to the DNA. Schematic representations of the coordination complexes formed by other metal ions bound in each site consistent with recent structural and spectroscopic studies are also shown., The α3 helices are indicated (see text for details).
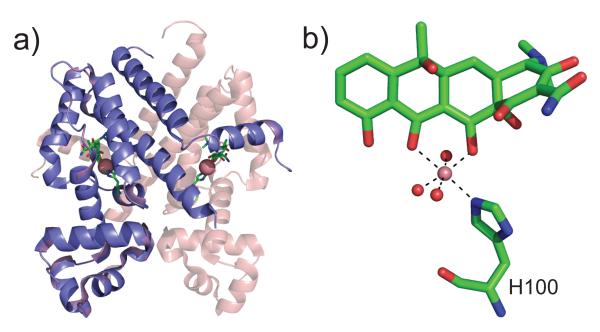
(a) Ribbon diagram of a class D Tetracycline repressor (TetR) bound to the antibiotic [Co7HTc]+ in a very deep pocket in the C-terminal core of regulatory domain (2VKE). The Co(II) ion is shaded red with the remainder of the antibiotic shown in stick representation. The core domain is at the top of the figure, while symmetry-related DNA-binding domains are shown at the bottom of the structure. (b) The conformation of tetracycline (7HTc) bound to the core domain, with Co(II) bound in an octahedral coordination geometry, as indicated. Water molecules are donated by red spheres while the imidazole Nε2 nitrogen of His100 donates the only Co(II)-side chain coordination bond. Mg(II) is the likely the biological metal ion and forms an isostructural complex.
References
-
- Lippard SJ, Berg JM. Principles of Bioinorganic Chemistry. University Science Books; Mill Valley, CA: 1994.
-
- Tottey S, Waldron KJ, Firbank SJ, Reale B, Bessant C, Sato K, Cheek TR, Gray J, Banfield MJ, Dennison C, Robinson NJ. Nature. 2008;455:1138. - PubMed
-
- Waldron KJ, Robinson NJ. Nat. Rev. Microbiol. 2009;7:25. - PubMed
-
- Duran RV, Hervas M, De La Rosa MA, Navarro JA. J. Biol. Chem. 2004;279:7229. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
