

Single molecule nanocontainers made porous using a bacterial toxin
- PMID: 19788247
- PMCID: PMC2761729
- DOI: 10.1021/ja9042356
Single molecule nanocontainers made porous using a bacterial toxin
Abstract
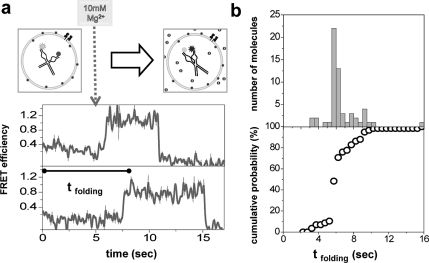
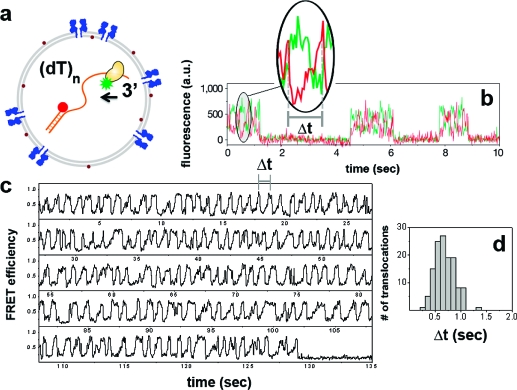
Encapsulation of a biological molecule or a molecular complex in a vesicle provides a means of biofriendly immobilization for single molecule studies and further enables new types of analysis if the vesicles are permeable. We previously reported on using DMPC (dimyristoylphosphatidylcholine) vesicles for realizing porous bioreactors. Here, we describe a different strategy for making porous vesicles using a bacterial pore-forming toxin, alpha-hemolysin. Using RNA folding as a test case, we demonstrate that protein-based pores can allow exchange of magnesium ions through the vesicle wall while keeping the RNA molecule inside. Flow measurements indicate that the encapsulated RNA molecules rapidly respond to the change in the outside buffer condition. The approach was further tested by coencapsulating a helicase protein and its single-stranded DNA track. The DNA translocation activity of E. coli Rep helicase inside vesicles was fueled by ATP provided outside the vesicle, and a dramatically higher number of translocation cycles could be observed due to the minuscule vesicle volume that facilitates rapid rebinding after dissociation. These pores are known to be stable over a wide range of experimental conditions, especially at various temperatures, which is not possible with the previous method using DMPC vesicles. Moreover, engineered mutants of the utilized toxin can potentially be exploited in the future applications.
Figures


References
-
- Joo C.; McKinney S. A.; Nakamura M.; Rasnik I.; Myong S.; Ha T. Cell 2006, 126, 515–27. - PubMed
-
- Joo C.; Balci H.; Ishitsuka Y.; Buranachai C.; Ha T. Annu. Rev. Biochem. 2008, 77, 51–76. - PubMed
-
- Xie X. S.; Choi P. J.; Li G. W.; Lee N. K.; Lia G. Annu. Rev. Biophys. 2008, 37, 417–44. - PubMed
-
- Yildiz A.; Forkey J. N.; McKinney S. A.; Ha T.; Goldman Y. E.; Selvin P. R. Science 2003, 300, 2061–5. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
