

Isotope labeling strategies for NMR studies of RNA
- PMID: 19789981
- PMCID: PMC2797625
- DOI: 10.1007/s10858-009-9375-2
Isotope labeling strategies for NMR studies of RNA
Abstract
The known biological functions of RNA have expanded in recent years and now include gene regulation, maintenance of sub-cellular structure, and catalysis, in addition to propagation of genetic information. As for proteins, RNA function is tightly correlated with structure. Unlike proteins, structural information for larger, biologically functional RNAs is relatively limited. NMR signal degeneracy, relaxation problems, and a paucity of long-range (1)H-(1)H dipolar contacts have limited the utility of traditional NMR approaches. Selective isotope labeling, including nucleotide-specific and segmental labeling strategies, may provide the best opportunities for obtaining structural information by NMR. Here we review methods that have been developed for preparing and purifying isotopically labeled RNAs, as well as NMR strategies that have been employed for signal assignment and structure determination.
Figures
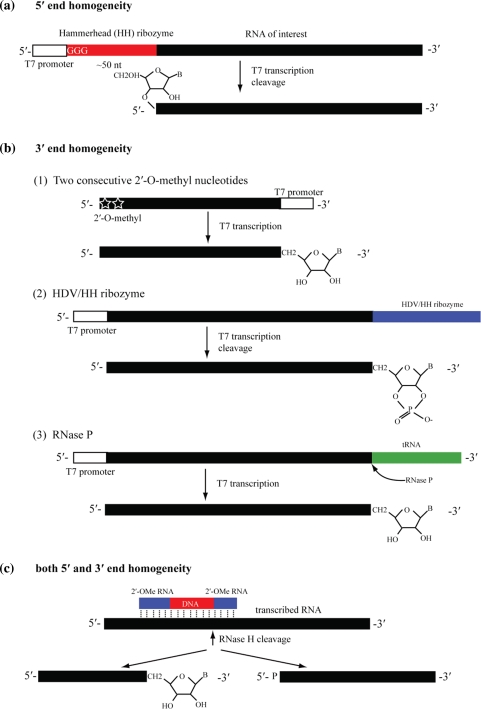
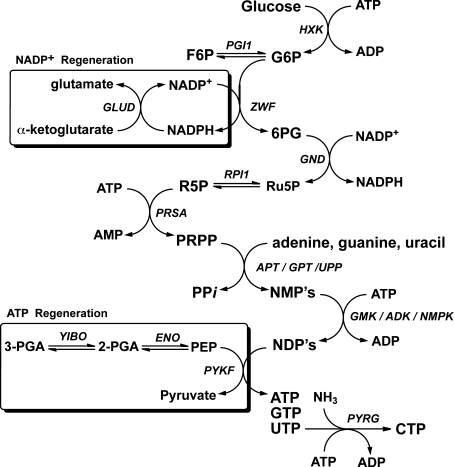
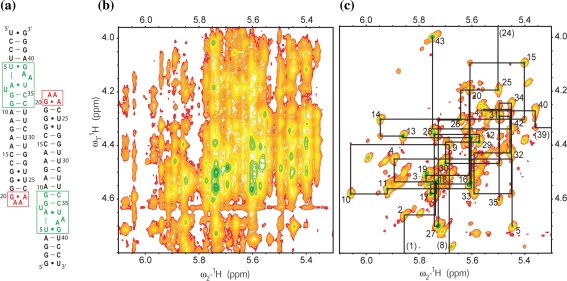
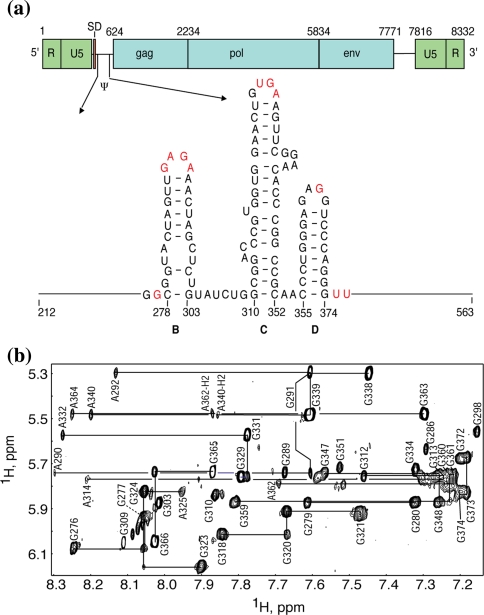
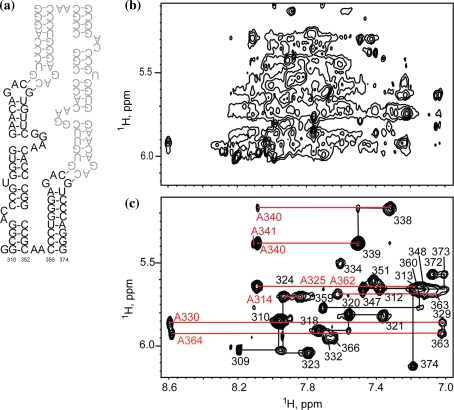

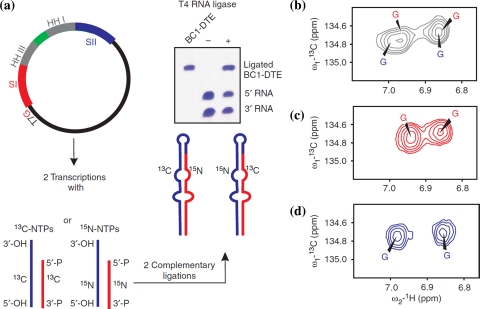
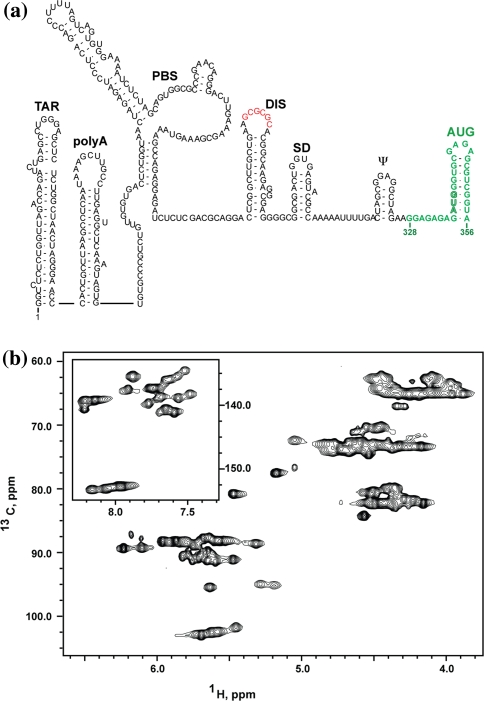
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
