

doi: 10.1101/gad.1836409.
Epub 2009 Sep 30.
The annealing helicase HARP protects stalled replication forks
Affiliations
- PMID: 19793864
- PMCID: PMC2764499
- DOI: 10.1101/gad.1836409
Item in Clipboard
The annealing helicase HARP protects stalled replication forks
Genes Dev.
.
Abstract
Mutations in HepA-related protein (HARP) are the only identified causes of Schimke immunoosseous dysplasia (SIOD). HARP has a unique annealing helicase activity in vitro, but the in vivo functional significance remains unknown. Here, we demonstrated that HARP is recruited to stalled replication forks via its direct interaction with Replication protein A (RPA). Cells with HARP depletion displayed increased spontaneous DNA damage and G2/M arrest, suggesting that HARP normally acts to stabilize stalled replication forks. Our data place the annealing helicase activity of HARP at replication forks and propose that SIOD syndrome may be caused by the destabilization of replication forks during cell proliferation.
Figures
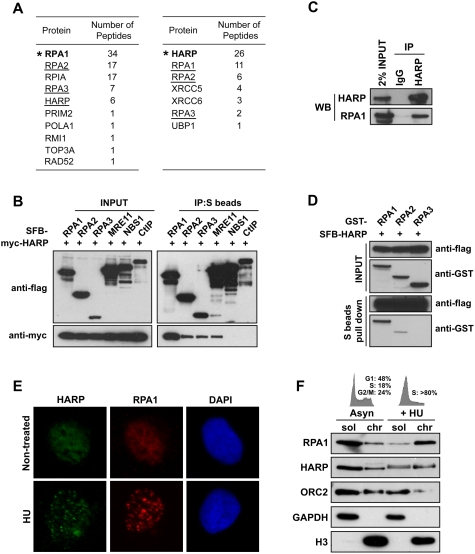
HARP associates with RPA complex and is recruited to stalled replication forks in response to replication stress. (A) TAP was performed using 293T cells stably expressing tagged RPA1 or HARP. The data from mass spectrometry analysis are shown in the tables. (B) The association of HARP with RPA complex was confirmed by coimmunoprecipitation with overexpressed proteins. 293T cells were transfected with plasmids encoding myc-tagged wild-type HARP together with plasmids encoding SFB-tagged RPA1, RPA2, RPA3, MRE11, NBS1, or CtIP. Cells were lysed 24 h after transfection. Coimmunoprecipitation was carried out using S-protein beads and immunoblotting was performed using antibodies as indicated. (C) Association of endogenous HARP with RPA1 in 293T cells was performed by coimmunoprecipitation using anti-HARP antibody. (D) HARP binds strongly to RPA1. The in vitro binding assay was performed using the baculovirus expression system. Sf9 cells were coinfected with baculoviruses expressing indicated constructs. Pull-down experiments were performed using S-protein beads and immunoblotting was carried out using indicated antibodies. (E) HARP localizes at stalled replication forks in response to replication stress. U2OS cells were mock-treated or treated with 5 mM HU for 6 h. Immunostaining experiments were performed using anti-HARP and anti-RPA1 antibodies. (F) HARP accumulates in chromatin fraction following HU treatment. Asynchronized or HU-treated U2OS cells were subjected to fractionation. Soluble (sol) and chromatin (chr) fractions were separated and immunoblotted with indicated antibodies. Cell cycle patterns are shown on top.
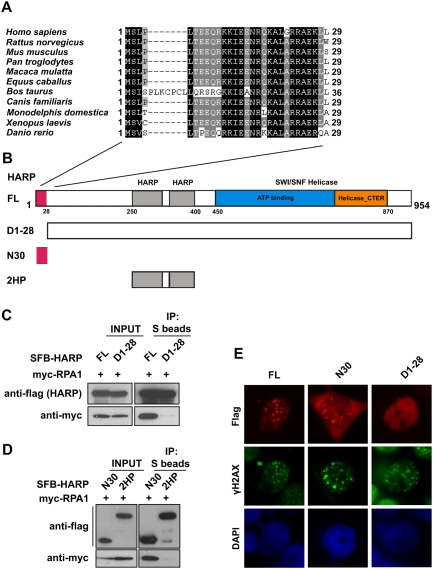
The conserved N terminus of HARP is required for RPA1 binding and its foci formation following replication stress. (A) Alignment of N-terminal sequences of HARP from different species. (B) Schematic representation of wild-type HARP and the mutants used in the following study. (C) The 28 amino acids at the N terminus of HARP are required for RPA1 binding. 293T cells were transfected with plasmids encoding myc-tagged RPA1 together with plasmids encoding SFB-tagged full-length HARP (FL) or the D1-28 mutant. Coimmunoprecipitation was carried out using S-protein beads and immunoblotting was performed using antibodies as indicated. (D) The N-terminal fragment (N30) of HARP is sufficient for RPA1 binding. Coimmunoprecipitation was performed similar to that described in C. The tandem HARP domain (2HP) was used as a control. (E) The conserved N terminus of HARP is responsible and sufficient for HARP foci formation following replication stress. 293T cells were transfected with plasmids encoding SFB-tagged full-length HARP (FL), the N30 fragment, or the D1-28 mutant. Immunostaining experiments were performed 6 h after HU treatment using indicated antibodies.
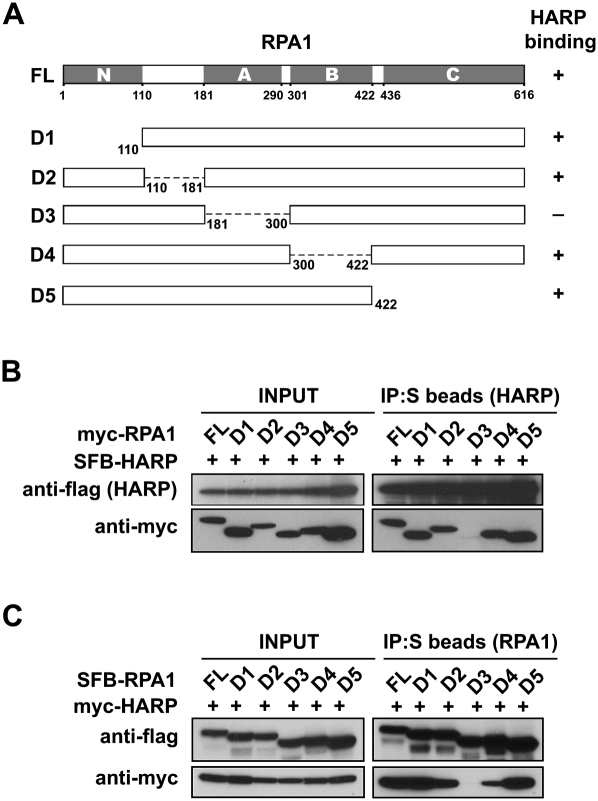
RPA1 associates with HARP through its ssDNA-binding domain A. (A) Schematic representation of full-length RPA1 and the mutants used in this study. Their ability to bind to HARP is indicated. (B,C) RPA1 with domain A deletion could not bind to HARP. 293T cells were transfected with plasmids encoding myc-tagged wild-type or mutant RPA1 together with plasmids encoding SFB-tagged HARP. Alternatively, 293T cells were transfected with plasmids encoding SFB-tagged wild-type or mutant RPA1 together with plasmids encoding myc-tagged HARP. Coimmunoprecipitation was carried out using S-protein beads and immunoblotting was performed using antibodies as indicated.
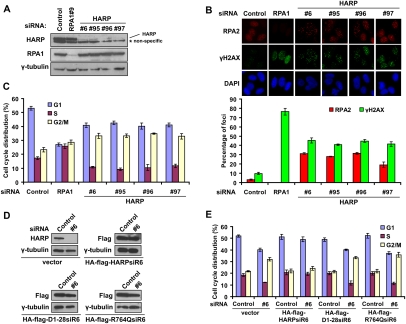
HARP depletion results in spontaneous DNA damage and G2/M cell cycle arrest. (A) Knockdown efficiency in U2OS cells transfected with indicated RPA1 and HARP siRNAs was confirmed by immunoblotting. (B) RPA and γH2AX foci are greatly increased in HARP-depleted cells. U2OS cells were transfected with indicated siRNAs. Seventy-two hours later, cells were subjected to immunostaining using indicated antibodies. The quantification of foci-positive cells was performed by counting a total of 200 cells per sample. (C) HARP depletion induces G2/M accumulation. Seventy-two hours after siRNAs transfection, U2OS cells were fixed and subjected to cell cycle analysis. The percentage of cells in different cell cycle phases was determined. Data were presented as mean ± SD from three different experiments. (D) U2OS derivative cell lines stably expressing siRNA-resistant wild-type HARP (HA-Flag-HARPsiR6), D1-28 mutants (HA-Flag-D1-28siR6), and R764Q mutants (HA-Flag-R764QsiR6) were generated. The endogenous and exogenous HARP expression was confirmed by immunoblotting using indicated antibodies and extracts were prepared from cells transfected with indicated siRNAs. (E) The D1-28 and R764Q mutants could not restore HARP function in vivo. Seventy-two hours after siRNA treatment, the indicated stable cells were fixed and subjected to cell cycle analysis. The percentage of cells in different cell cycle phases was determined. Data are presented as mean ± SD from three different experiments.
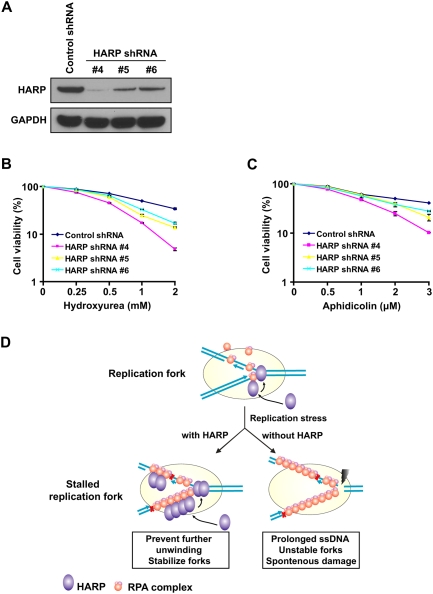
HARP insufficiency causes hypersensitivity to replication stress. (A) Knockdown efficiency of HARP in HeLa cells stably expressing the indicated shRNA was confirmed by immunoblotting. (B) Survival curves in response to increasing doses of HU for indicated cell lines. (C) Survival curves in response to increasing doses of aphidicolin for indicated cell lines. Cell survival assays were performed according to the Materials and Methods. Data are presented as mean ± SD from three different experiments. (D) A proposed model of HARP function at replication forks. See the text for details.
Comment in
-
HARPing on about the DNA damage response during replication.Genes Dev. 2009 Oct 15;23(20):2359-65. doi: 10.1101/gad.1860609. Genes Dev. 2009. PMID: 19833762 Free PMC article.
References
-
- Binz SK, Sheehan AM, Wold MS. Replication protein A phosphorylation and the cellular response to DNA damage. DNA Repair (Amst) 2004;3:1015–1024. - PubMed
-
- Boerkoel CF, O'Neill S, Andre JL, Benke PJ, Bogdanovic R, Bulla M, Burguet A, Cockfield S, Cordeiro I, Ehrich JH, et al. Manifestations and treatment of Schimke immuno-osseous dysplasia: 14 new cases and a review of the literature. Eur J Pediatr. 2000;159:1–7. - PubMed
-
- Boerkoel CF, Takashima H, John J, Yan J, Stankiewicz P, Rosenbarker L, Andre JL, Bogdanovic R, Burguet A, Cockfield S, et al. Mutant chromatin remodeling protein SMARCAL1 causes Schimke immuno-osseous dysplasia. Nat Genet. 2002;30:215–220. - PubMed
-
- Brosh RM, Jr, Li JL, Kenny MK, Karow JK, Cooper MP, Kureekattil RP, Hickson ID, Bohr VA. Replication protein A physically interacts with the Bloom's syndrome protein and stimulates its helicase activity. J Biol Chem. 2000;275:23500–23508. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources