

Talin 1 and 2 are required for myoblast fusion, sarcomere assembly and the maintenance of myotendinous junctions
- PMID: 19793892
- PMCID: PMC2761109
- DOI: 10.1242/dev.035857
Talin 1 and 2 are required for myoblast fusion, sarcomere assembly and the maintenance of myotendinous junctions
Abstract
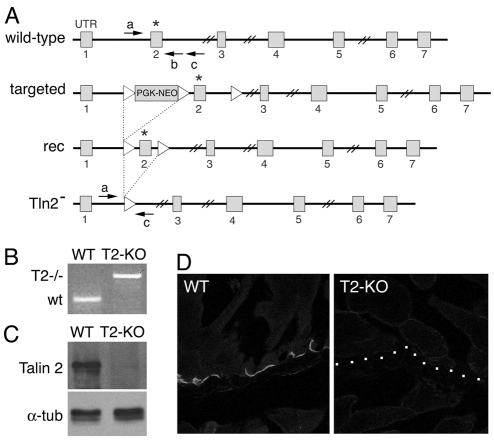
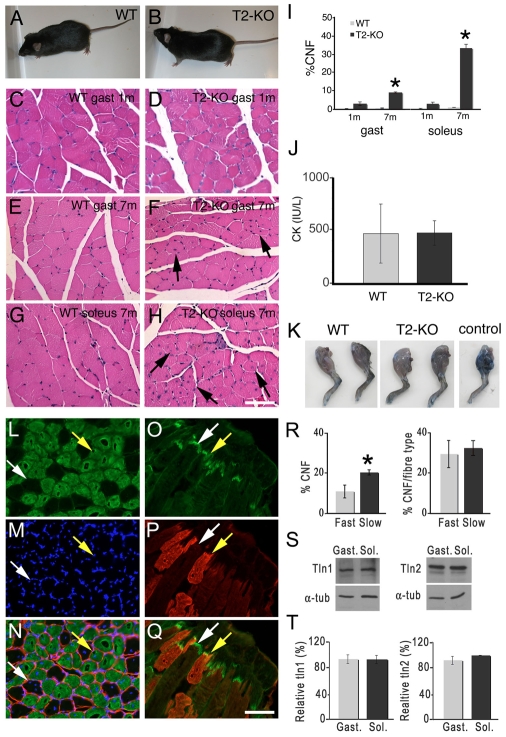
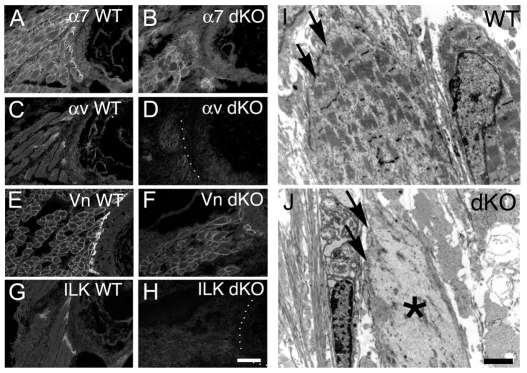
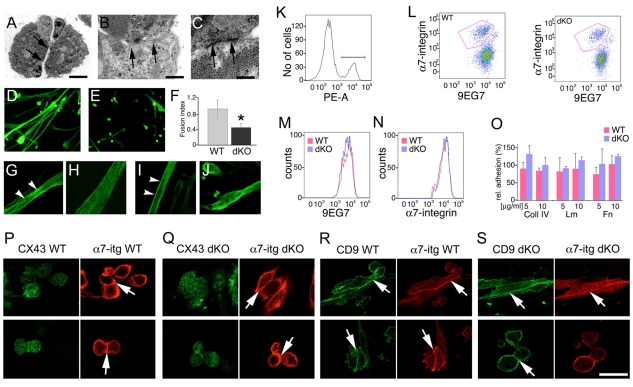
Talin 1 and 2 connect integrins to the actin cytoskeleton and regulate the affinity of integrins for ligands. In skeletal muscle, talin 1 regulates the stability of myotendinous junctions (MTJs), but the function of talin 2 in skeletal muscle is not known. Here we show that MTJ integrity is affected in talin 2-deficient mice. Concomitant ablation of talin 1 and 2 leads to defects in myoblast fusion and sarcomere assembly, resembling defects in muscle lacking beta1 integrins. Talin 1/2-deficient myoblasts express functionally active beta1 integrins, suggesting that defects in muscle development are not primarily caused by defects in ligand binding, but rather by disruptions of the interaction of integrins with the cytoskeleton. Consistent with this finding, assembly of integrin adhesion complexes is perturbed in the remaining muscle fibers of talin 1/2-deficient mice. We conclude that talin 1 and 2 are crucial for skeletal muscle development, where they regulate myoblast fusion, sarcomere assembly and the maintenance of MTJs.
Figures



References
-
- Araya, R., Eckardt, D., Maxeiner, S., Kruger, O., Theis, M., Willecke, K. and Saez, J. C. (2005). Expression of connexins during differentiation and regeneration of skeletal muscle: functional relevance of connexin43. J. Cell Sci. 118, 27-37. - PubMed
-
- Balaban, N. Q., Schwarz, U. S., Riveline, D., Goichberg, P., Tzur, G., Sabanay, I., Mahalu, D., Safran, S., Bershadsky, A., Addadi, L. et al. (2001). Force and focal adhesion assembly: a close relationship studied using elastic micropatterned substrates. Nat. Cell Biol. 3, 466-472. - PubMed
-
- Bazzoni, G., Shih, D. T., Buck, C. A. and Hemler, M. E. (1995). Monoclonal antibody 9EG7 defines a novel beta 1 integrin epitope induced by soluble ligand and manganese, but inhibited by calcium. J. Biol. Chem. 270, 25570-25577. - PubMed
-
- Belkin, A. M., Zhidkova, N. I., Balzac, F., Altruda, F., Tomatis, D., Maier, A., Tarone, G., Koteliansky, V. E. and Burridge, K. (1996). Beta 1D integrin displaces the beta 1A isoform in striated muscles: localization at junctional structures and signaling potential in nonmuscle cells. J. Cell Biol. 132, 211-226. - PMC - PubMed
-
- Belkin, A. M., Retta, S. F., Pletjushkina, O. Y., Balzac, F., Silengo, L., Fassler, R., Koteliansky, V. E., Burridge, K. and Tarone, G. (1997). Muscle beta1D integrin reinforces the cytoskeleton-matrix link: modulation of integrin adhesive function by alternative splicing. J. Cell Biol. 139, 1583-1595. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
