

Requirement of cellular DDX3 for hepatitis C virus replication is unrelated to its interaction with the viral core protein
- PMID: 19793905
- PMCID: PMC2885062
- DOI: 10.1099/vir.0.015909-0
Requirement of cellular DDX3 for hepatitis C virus replication is unrelated to its interaction with the viral core protein
Abstract
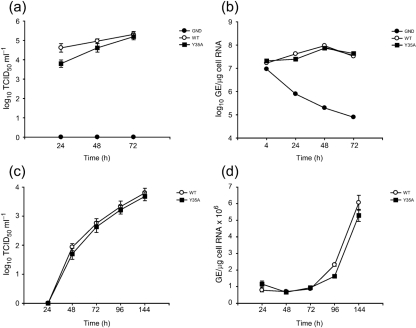
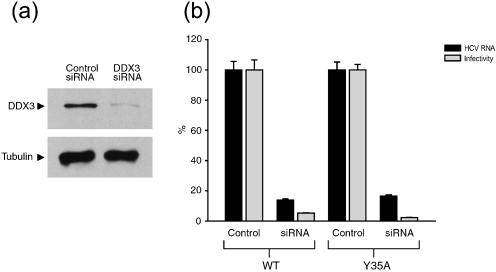
The cellular DEAD-box protein DDX3 was recently shown to be essential for hepatitis C virus (HCV) replication. Prior to that, we had reported that HCV core binds to DDX3 in yeast-two hybrid and transient transfection assays. Here, we confirm by co-immunoprecipitation that this interaction occurs in cells replicating the JFH1 virus. Consistent with this result, immunofluorescence staining of infected cells revealed a dramatic redistribution of cytoplasmic DDX3 by core protein to the virus assembly sites around lipid droplets. Given this close association of DDX3 with core and lipid droplets, and its involvement in virus replication, we investigated the importance of this host factor in the virus life cycle. Mutagenesis studies located a single amino acid in the N-terminal domain of JFH1 core that when changed to alanine significantly abrogated this interaction. Surprisingly, this mutation did not alter infectious virus production and RNA replication, indicating that the core-DDX3 interaction is dispensable in the HCV life cycle. Consistent with previous studies, siRNA-led knockdown of DDX3 lowered virus production and RNA replication levels of both WT JFH1 and the mutant virus unable to bind DDX3. Thus, our study shows for the first time that the requirement of DDX3 for HCV replication is unrelated to its interaction with the viral core protein.
Figures
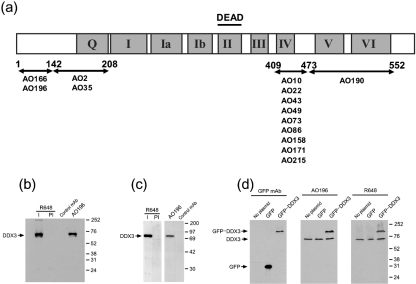
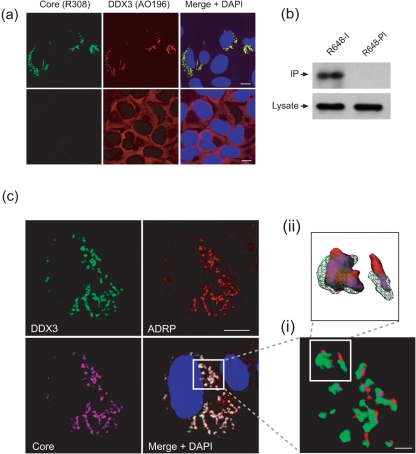

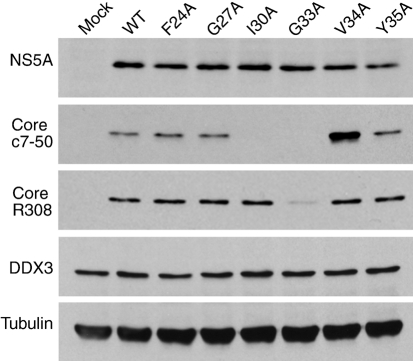
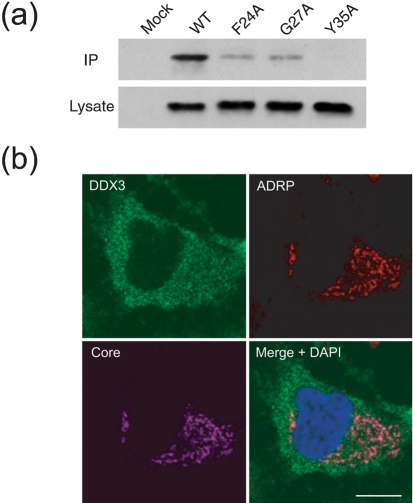
References
-
- Barba, G., Harper, F., Harada, T., Kohara, M., Goulinet, S., Matsuura, Y., Eder, G., Schaff, Z., Chapman, M. J. & other authors (1997). Hepatitis C virus core protein shows a cytoplasmic localization and associates to cellular lipid storage droplets. Proc Natl Acad Sci U S A 94, 1200–1205. - PMC - PubMed
-
- Blight, K. J., Kolykhalov, A. A. & Rice, C. M. (2000). Efficient initiation of HCV RNA replication in cell culture. Science 290, 1972–1974. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources