

Control of NMDA receptor function by the NR2 subunit amino-terminal domain
- PMID: 19793963
- PMCID: PMC2776059
- DOI: 10.1523/JNEUROSCI.1365-09.2009
Control of NMDA receptor function by the NR2 subunit amino-terminal domain
Abstract
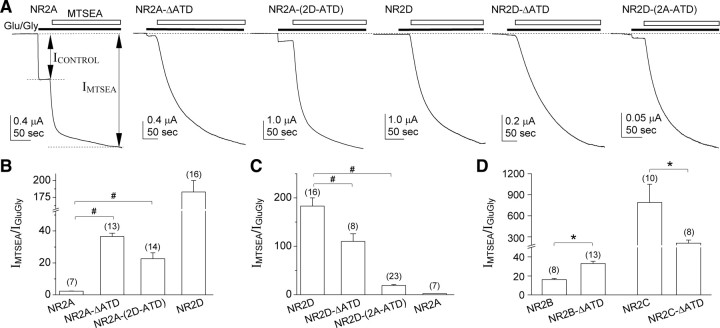
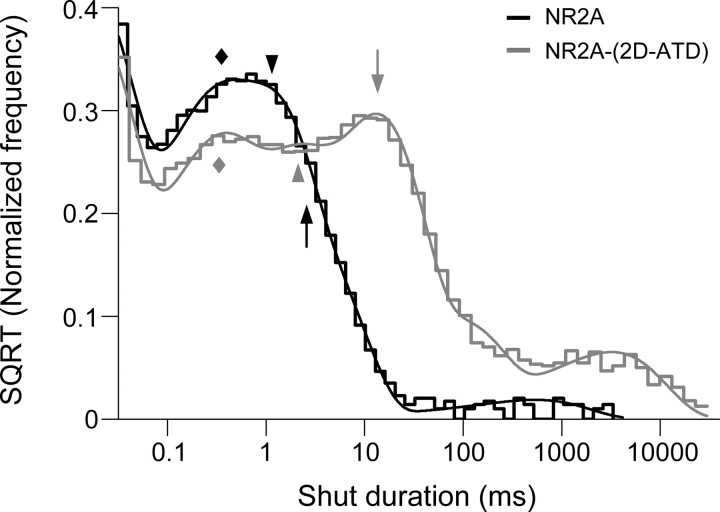
NMDA receptors comprised of different NR2 subunits exhibit strikingly unique biophysical and pharmacological properties. Here, we report that the extracellular amino-terminal domain (ATD) of the NR2 subunit controls pharmacological and kinetic properties of recombinant NMDA receptors, such as agonist potency, deactivation time course, open probability (P(OPEN)), and mean open/shut duration. Using ATD deletion mutants of NR2A, NR2B, NR2C, NR2D, and chimeras of NR2A and NR2D with interchanged ATD [NR2A-(2D-ATD) and NR2D-(2A-ATD)], we show that the ATD contributes to the low glutamate potency of NR2A-containing NMDA receptors and the high glutamate potency of NR2D-containing receptors. The ATD influences the deactivation time courses of NMDA receptors, as removal of the ATD from NR2A slows the deactivation rate, while removal of the ATD from NR2B, NR2C and NR2D accelerates the deactivation rate. Open probability also is influenced by the ATD. Removal of the ATD from NR2A or replacement of the NR2A-ATD with that of NR2D decreases P(OPEN) in single-channel recordings from outside-out patches of HEK 293 cells. In contrast, deletion of the ATD from NR2D or replacement of the NR2D ATD with that of NR2A increases P(OPEN) and mean open duration. These data demonstrate the modular nature of NMDA receptors, and show that the ATD of the different NR2 subunits plays an important role in fine-tuning the functional properties of the individual NMDA receptor subtypes.
Figures








References
-
- Béhé P, Colquhoun D, Wyllie DJ. Activation of Single AMPA- and NMDA-Type Glutamate-Receptor Channels. In: Jonas P, Monyer H, editors. Ionotropic glutamate receptors in the CNS. Springer; 1999. pp. 175–218.
-
- Choi YB, Lipton SA. Identification and mechanism of action of two histidine residues underlying high-affinity Zn2+ inhibition of the NMDA receptor. Neuron. 1999;23:171–180. - PubMed
-
- Colquhoun D, Hawkes AG. Stochastic properties of ion channel openings and bursts in a membrane patch that contains two channels: evidence concerning the number of channels present when a record containing only single openings is observed. Proc R Soc Lond B Biol Sci. 1990;240:453–477. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous