

Behavioral deficits and subregion-specific suppression of LTP in mice expressing a population of mutant NMDA receptors throughout the hippocampus
- PMID: 19794189
- PMCID: PMC2769167
- DOI: 10.1101/lm.1316909
Behavioral deficits and subregion-specific suppression of LTP in mice expressing a population of mutant NMDA receptors throughout the hippocampus
Abstract
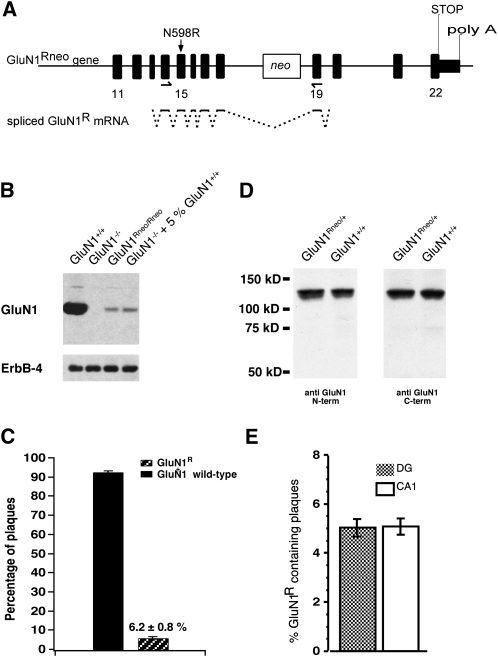
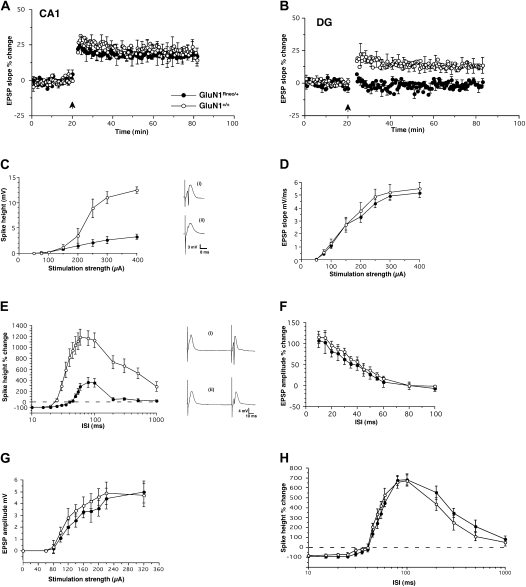
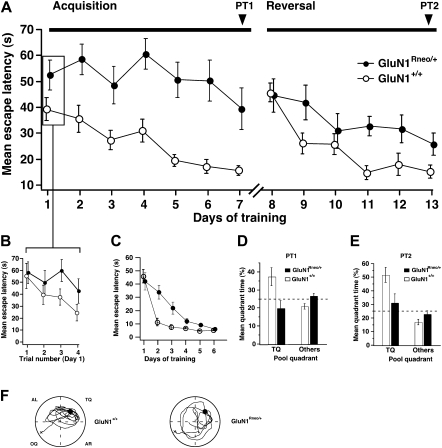
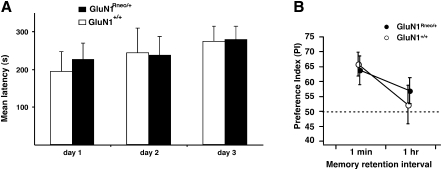
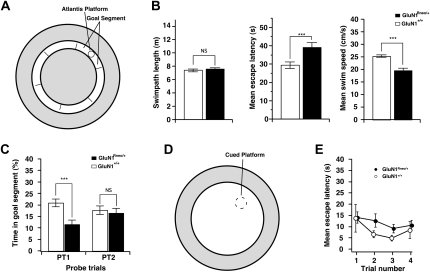
The NMDA receptor (NMDAR) subunit GluN1 is an obligatory component of NMDARs without a known functional homolog and is expressed in almost every neuronal cell type. The NMDAR system is a coincidence detector with critical roles in spatial learning and synaptic plasticity. Its coincidence detection property is crucial for the induction of hippocampal long-term potentiation (LTP). We have generated a mutant mouse model expressing a hypomorph of the Grin1(N598R) allele, which leads to a minority (about 10%) of coincidence detection-impaired NMDARs. Surprisingly, these animals revealed specific functional changes in the dentate gyrus (DG) of the hippocampal formation. Early LTP was expressed normally in area CA1 in vivo, but was completely suppressed at perforant path-granule cell synapses in the DG. In addition, there was a pronounced reduction in the amplitude of the evoked population spike in the DG. These specific changes were accompanied by behavioral impairments in spatial recognition, spatial learning, reversal learning, and retention. Our data show that minor changes in GluN1-dependent NMDAR physiology can cause dramatic consequences in synaptic signaling in a subregion-specific fashion despite the nonredundant nature of the GluN1 gene and its global expression.
Figures
Similar articles
-
Interneuronal NMDA receptors regulate long-term depression and motor learning in the cerebellum.J Physiol. 2019 Feb;597(3):903-920. doi: 10.1113/JP276794. Epub 2018 Nov 24. J Physiol. 2019. PMID: 30382582 Free PMC article.
-
Impaired spatial working memory but spared spatial reference memory following functional loss of NMDA receptors in the dentate gyrus.Eur J Neurosci. 2007 Feb;25(3):837-46. doi: 10.1111/j.1460-9568.2007.05312.x. Eur J Neurosci. 2007. PMID: 17313573 Free PMC article.
-
SynGAP regulates ERK/MAPK signaling, synaptic plasticity, and learning in the complex with postsynaptic density 95 and NMDA receptor.J Neurosci. 2002 Nov 15;22(22):9721-32. doi: 10.1523/JNEUROSCI.22-22-09721.2002. J Neurosci. 2002. PMID: 12427827 Free PMC article.
-
Hippocampal long-term synaptic plasticity and signal amplification of NMDA receptors.Crit Rev Neurobiol. 2006;18(1-2):71-84. doi: 10.1615/critrevneurobiol.v18.i1-2.80. Crit Rev Neurobiol. 2006. PMID: 17725510 Review.
-
[Neuronal plasticity associated with learning and epileptic seizures: LTP and KIP].Seishin Shinkeigaku Zasshi. 2001;103(10):866-81. Seishin Shinkeigaku Zasshi. 2001. PMID: 11797444 Review. Japanese.
Cited by
-
Examining Neurosteroid-Analogue Therapy in the Preterm Neonate For Promoting Hippocampal Neurodevelopment.Front Physiol. 2022 Apr 19;13:871265. doi: 10.3389/fphys.2022.871265. eCollection 2022. Front Physiol. 2022. PMID: 35514343 Free PMC article.
-
Neuroligin-3 Regulates Excitatory Synaptic Transmission and EPSP-Spike Coupling in the Dentate Gyrus In Vivo.Mol Neurobiol. 2022 Feb;59(2):1098-1111. doi: 10.1007/s12035-021-02663-9. Epub 2021 Nov 29. Mol Neurobiol. 2022. PMID: 34845591 Free PMC article.
-
Very low density lipoprotein receptor regulates dendritic spine formation in a RasGRF1/CaMKII dependent manner.Biochim Biophys Acta. 2015 May;1853(5):904-17. doi: 10.1016/j.bbamcr.2015.01.015. Epub 2015 Jan 31. Biochim Biophys Acta. 2015. PMID: 25644714 Free PMC article.
-
Optical imaging of plastic changes induced by fear conditioning in the auditory cortex.Cogn Neurodyn. 2012 Feb;6(1):1-10. doi: 10.1007/s11571-011-9173-x. Epub 2011 Aug 30. Cogn Neurodyn. 2012. PMID: 23372615 Free PMC article.
-
β-Estradiol unmasks metabotropic receptor-mediated metaplasticity of NMDA receptor transmission in the female rat dentate gyrus.Psychoneuroendocrinology. 2012 Nov;37(11):1845-54. doi: 10.1016/j.psyneuen.2012.03.023. Epub 2012 Apr 27. Psychoneuroendocrinology. 2012. PMID: 22541715 Free PMC article.
References
-
- Behe P, Stern P, Wyllie DJ, Nassar M, Schoepfer R, Colquhoun D. Determination of NMDA NR1 subunit copy number in recombinant NMDA receptors. Proc R Soc Lond B Biol Sci. 1995;262:205–213. - PubMed
-
- Bliss TV, Collingridge GL. A synaptic model of memory: Long-term potentiation in the hippocampus. Nature. 1993;361:31–39. - PubMed
-
- Bliss T, Errington M, Fransen E, Godfraind JM, Kauer JA, Kooy RF, Maness PF, Furley AJ. Long-term potentiation in mice lacking the neural cell adhesion molecule L1. Curr Biol. 2000;10:1607–1610. - PubMed
-
- Brun VH, Otnass MK, Molden S, Steffenach HA, Witter MP, Moser MB, Moser EI. Place cells and place recognition maintained by direct entorhinal-hippocampal circuitry. Science. 2002;296:2243–2246. - PubMed
-
- Burnashev N, Schoepfer R, Monyer H, Ruppersberg JP, Gunther W, Seeburg PH, Sakmann B. Control by asparagine residues of calcium permeability and magnesium blockade in the NMDA receptor. Science. 1992;257:1415–1419. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous