

The multilevel and dynamic interplay between plant and pathogen
- PMID: 19794843
- PMCID: PMC2664487
- DOI: 10.4161/psb.4.4.8155
The multilevel and dynamic interplay between plant and pathogen
Retraction in
-
Retraction of : "The multilevel and dynamic interplay between plant and pathogen".Plant Signal Behav. 2009 Jun;4(6):568. doi: 10.4161/psb.4.6.8867. Epub 2009 Jun 28. Plant Signal Behav. 2009. PMID: 19816134 Free PMC article. No abstract available.
Abstract
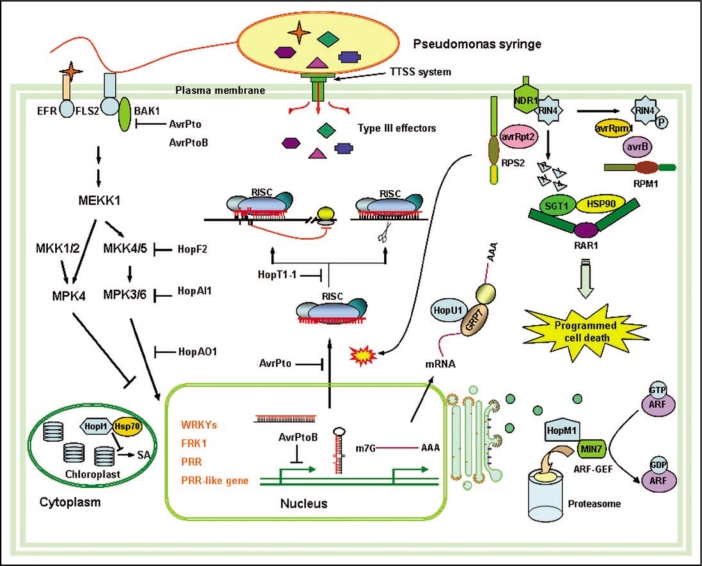
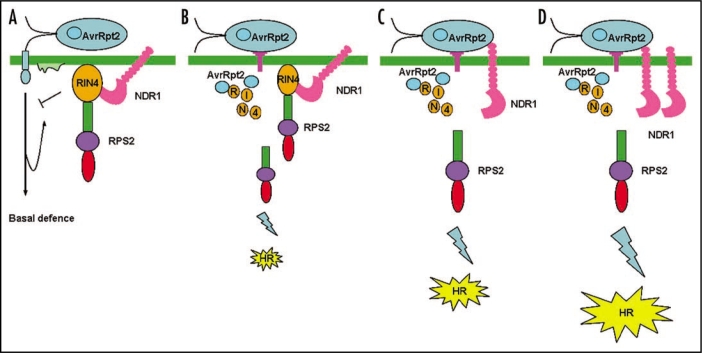
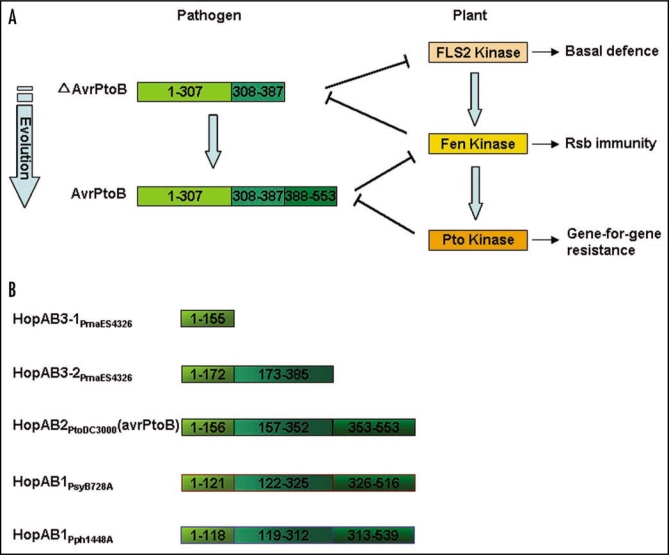
Phytopathogens invade into plant apoplast and proliferate by assimilating nutrition from plant cells. Plants depend on sophisticated defensive strategies to resist this invasion. Therefore, pathogenic disease and plant disease resistance are two opposite phases. Fascinating molecular mechanisms uncovered that interactions between plant and pathogen are multilevel and dynamic processes. On one side, plant immunity system contains multiple layers mainly including the perception of common pathogen- associated molecular patterns (PAMPs) using distinct cell-surface pattern-recognition receptors (PRRs) to activate intracellular signaling pathways for broad-spectrum immunity, and the recognition of pathogen virulence proteins by the specific intracellular disease resistance (R) proteins for cultivar-specific immunity. On the opposite side, the bacterial pathogens employ virulence factors, such as phytotoxin and type III effectors (T3SEs) to interfere with the host immunity in different levels. Meanwhile, natural selection drives plants and pathogens to evolve new strategies to confront with each other constantly. The present review highlights recent insights about Arabidopsis immunity and mechanisms for Pseudomonas syringae to counteract this immunity to give a full understanding of plant-pathogen interactions.
Keywords: Pseudomonas syringae; arabidopsis; evolution; plant-pathogen interactions; type III effector.
Figures
References
-
- Chisholm ST, Coaker G, Day B, Staskawicz BJ. Host-microbe interactions: Shaping the evolution of the plant immune response. Cell. 2006;124:803–814. - PubMed
-
- Jones JD, Dangl JL. The plant immune system. Nature. 2006;444:323–329. - PubMed
-
- Nürnberger T, Kemmerling B. Receptor protein kinases—pattern recognition receptors in plant immunity. Trends Plant Sci. 2006;11:519–522. - PubMed
-
- Ligterink W, Kroj T, zur Nieden U, Hirt H, Scheel D. Receptor-mediated activation of a MAP kinase in pathogen defence of plants. Science. 1997;276:2054–2057. - PubMed
-
- Asai T, Tena G, Plotnikova J, Willmann MR, Chiu WL, Gomez-Gomez L, et al. MAP kinase signaling cascade in Arabidopsis innate immunity. Nature. 2002;415:977–983. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources