

Histone H1 subtypes differentially modulate chromatin condensation without preventing ATP-dependent remodeling by SWI/SNF or NURF
- PMID: 19794910
- PMCID: PMC2748705
- DOI: 10.1371/journal.pone.0007243
Histone H1 subtypes differentially modulate chromatin condensation without preventing ATP-dependent remodeling by SWI/SNF or NURF
Abstract
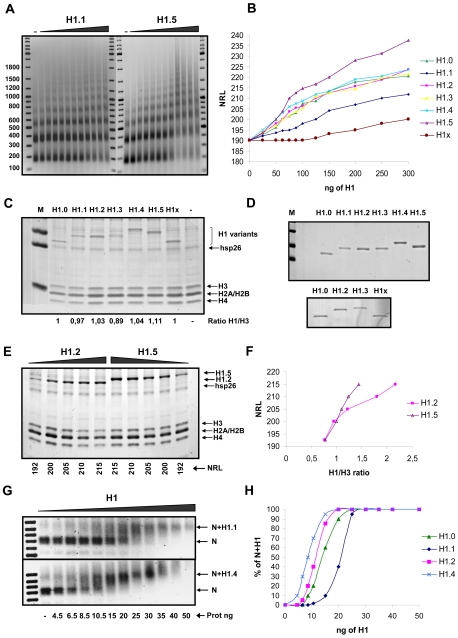
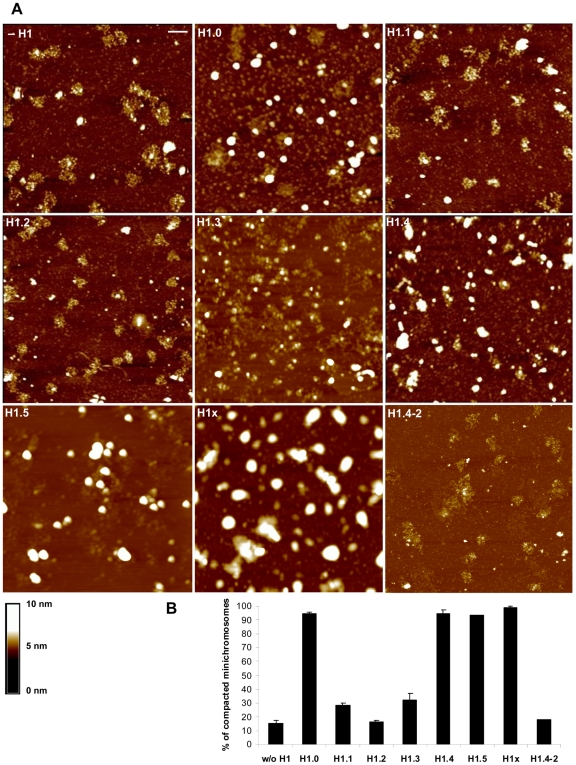
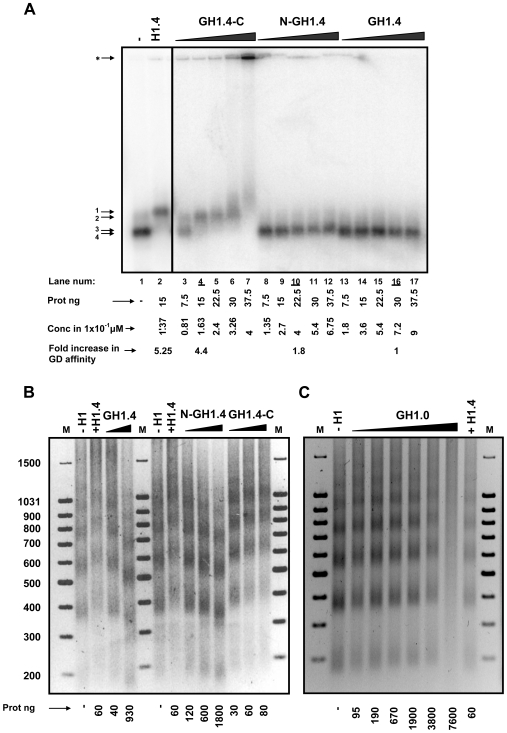
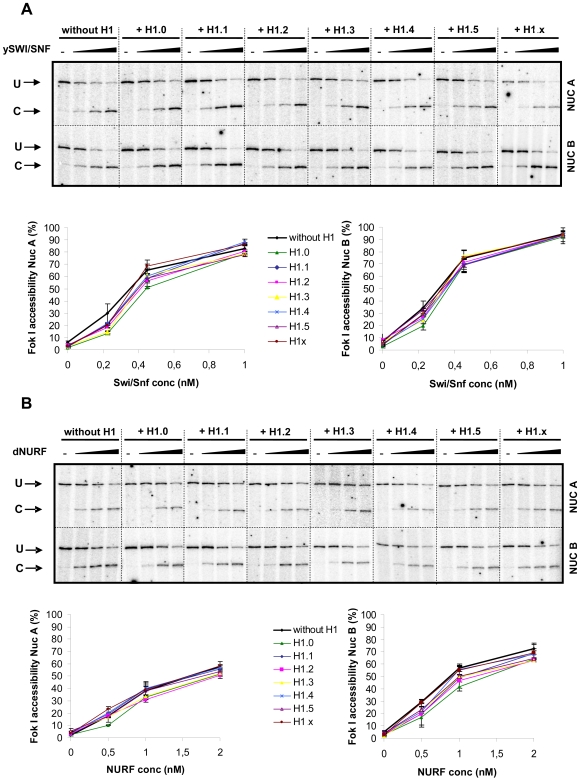
Although ubiquitously present in chromatin, the function of the linker histone subtypes is partly unknown and contradictory studies on their properties have been published. To explore whether the various H1 subtypes have a differential role in the organization and dynamics of chromatin we have incorporated all of the somatic human H1 subtypes into minichromosomes and compared their influence on nucleosome spacing, chromatin compaction and ATP-dependent remodeling. H1 subtypes exhibit different affinities for chromatin and different abilities to promote chromatin condensation, as studied with the Atomic Force Microscope. According to this criterion, H1 subtypes can be classified as weak condensers (H1.1 and H1.2), intermediate condensers (H1.3) and strong condensers (H1.0, H1.4, H1.5 and H1x). The variable C-terminal domain is required for nucleosome spacing by H1.4 and is likely responsible for the chromatin condensation properties of the various subtypes, as shown using chimeras between H1.4 and H1.2. In contrast to previous reports with isolated nucleosomes or linear nucleosomal arrays, linker histones at a ratio of one per nucleosome do not preclude remodeling of minichromosomes by yeast SWI/SNF or Drosophila NURF. We hypothesize that the linker histone subtypes are differential organizers of chromatin, rather than general repressors.
Conflict of interest statement
Figures

References
-
- Luger K, Mader AW, Richmond RK, Sargent DF, Richmond TJ. Crystal structure of the nucleosome core particle at 2.8 A resolution. Nature. 1997;389:251–260. - PubMed
-
- Ramakrishnan V. Histone structure and the organization of the nucleosome. Annu Rev Biophys Biomol Struct. 1997;26:83–112. - PubMed
-
- Huynh VA, Robinson PJ, Rhodes D. A method for the in vitro reconstitution of a defined “30 nm” chromatin fibre containing stoichiometric amounts of the linker histone. J Mol Biol. 2005;345:957–968. - PubMed
-
- Butler PJ, Thomas JO. Changes in chromatin folding in solution. J Mol Biol. 1980;140:505–529. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
