

Proteomic comparison of four Eimeria tenella life-cycle stages: unsporulated oocyst, sporulated oocyst, sporozoite and second-generation merozoite
- PMID: 19795439
- PMCID: PMC2947549
- DOI: 10.1002/pmic.200900305
Proteomic comparison of four Eimeria tenella life-cycle stages: unsporulated oocyst, sporulated oocyst, sporozoite and second-generation merozoite
Abstract
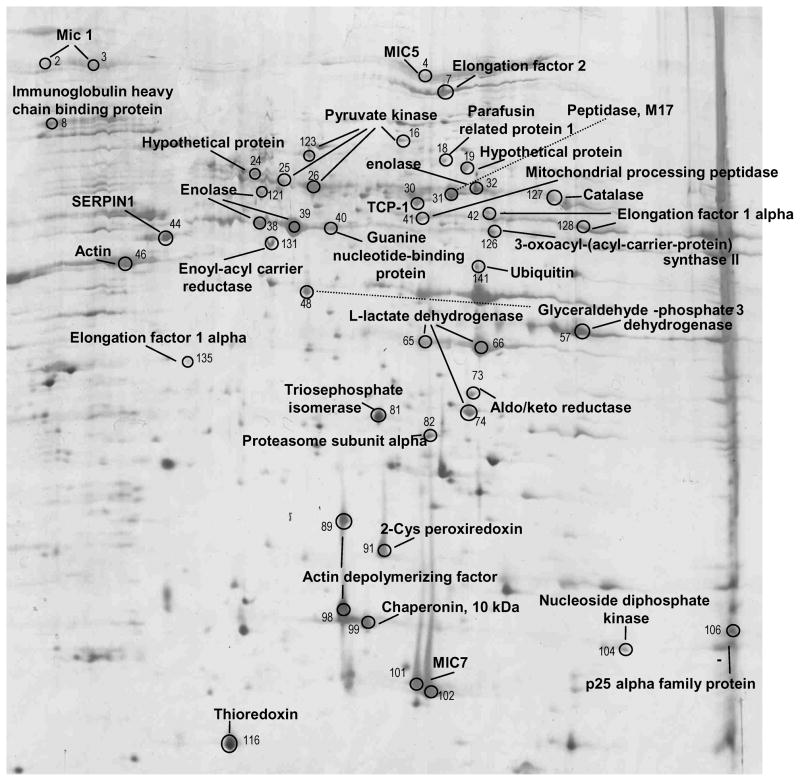
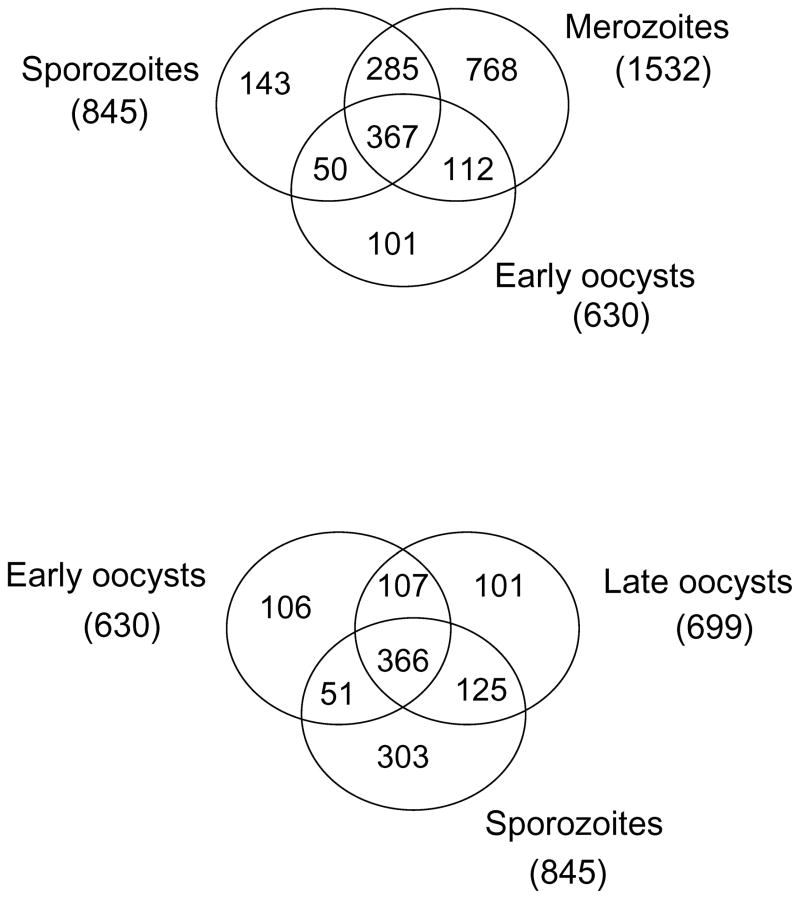
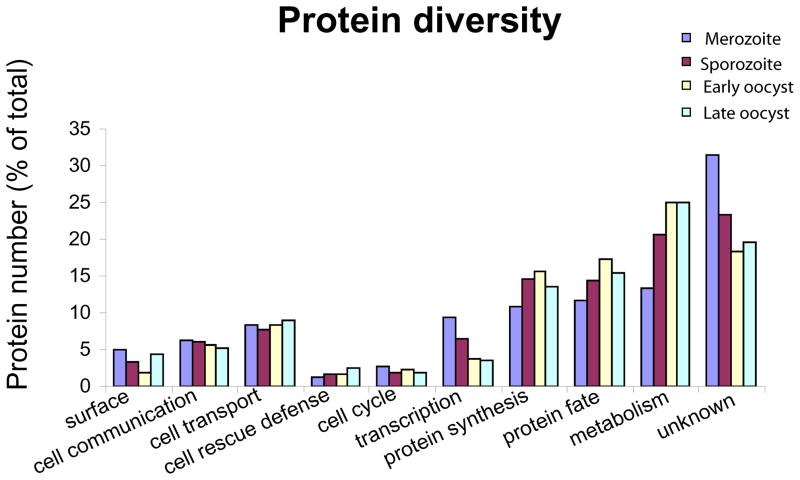
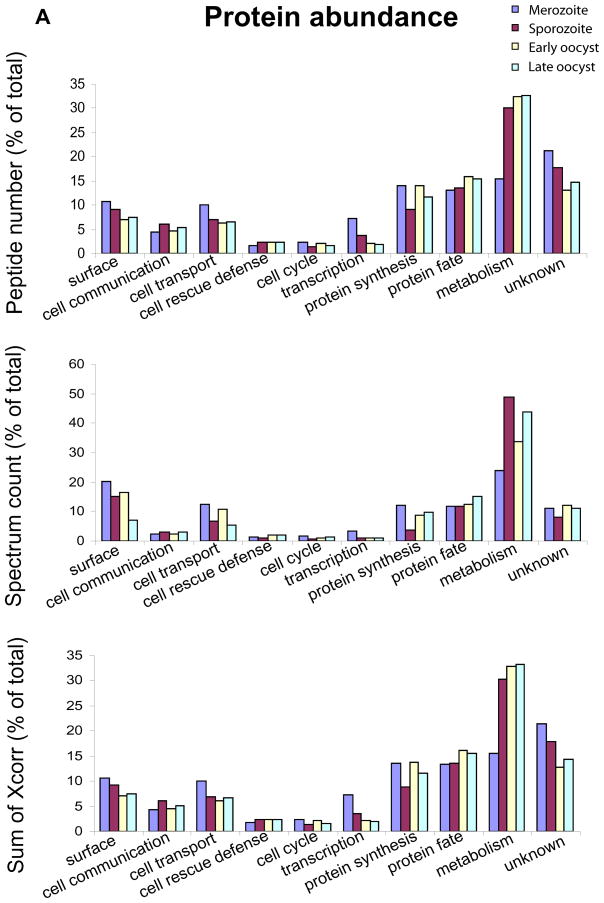
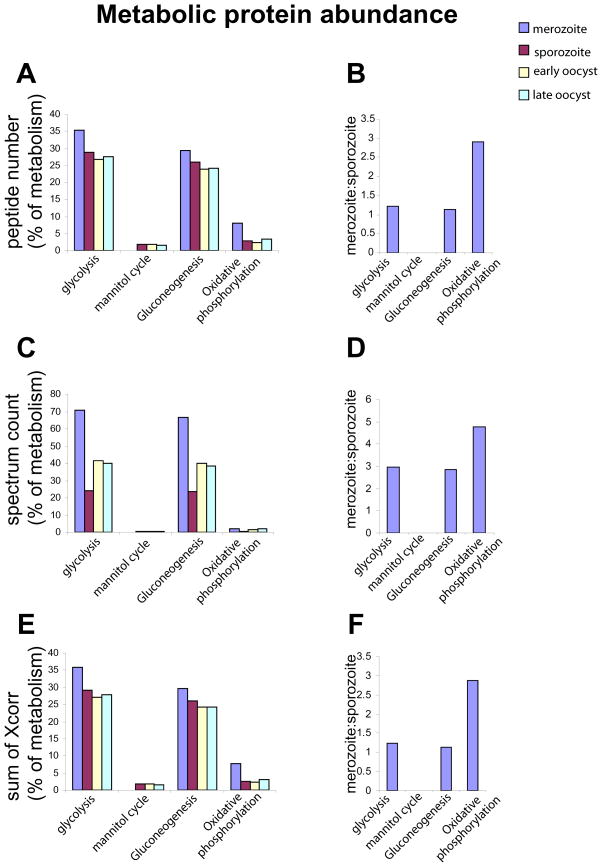
We report the proteomes of four life-cycle stages of the Apicomplexan parasite Eimeria tenella. A total of 1868 proteins were identified, with 630, 699, 845 and 1532 found in early oocysts (unsporulated), late oocysts (sporulated), sporozoites and second-generation merozoites, respectively. A multidimensional protein identification technology shotgun approach identified 812 sporozoites, 1528 merozoites and all of the oocyst proteins, whereas 2-D gel proteomics identified 230 sporozoites and 98 merozoite proteins. Comparing the invasive stages, we find moving junction components RON2 in both, whereas AMA-1 and RON4 are found only in merozoites and AMA-2 and RON5 are only found in sporozoites, suggesting stage-specific moving junction proteins. During early oocyst to sporozoite development, refractile body and most "glideosome" proteins are found throughout, whereas microneme and most rhoptry proteins are only found after sporulation. Quantitative analysis indicates glycolysis and gluconeogenesis are the most abundant metabolic groups detected in all stages. The mannitol cycle "off shoot" of glycolysis was not detected in merozoites but was well represented in the other stages. However, in merozoites we find more protein associated with oxidative phosphorylation, suggesting a metabolic shift mobilising greater energy production. We find a greater abundance of protein linked to transcription, protein synthesis and cell cycle in merozoites than in sporozoites, which may be residual protein from the preceding massive replication during schizogony.
Figures

References
-
- Shirley MW, Smith AL, Tomley FM. The biology of avian Eimeria with an emphasis on their control by vaccination. Adv Parasitol. 2005;60:285–330. - PubMed
-
- Ferguson DJ, Belli SI, Smith NC, Wallach MG. The development of the macrogamete and oocyst wall in Eimeria maxima: immuno-light and electron microscopy. Int J Parasitol. 2003;33:1329–1340. - PubMed
-
- Ryan R, Shirley M, Tomley F. Mapping and expression of microneme genes in Eimeria tenella. Int J Parasitol. 2000;30:1493–1499. - PubMed
-
- Rose ME, Lawn AM, Millard BJ. The effect of immunity on the early events in the life-cycle of Eimeria tenella in the caecal mucosa of the chicken. Parasitology. 1984;88:199–210. - PubMed
-
- Lawn AM, Rose ME. Mucosal transport of Eimeria tenella in the cecum of the chicken. J Parasitol. 1982;68:1117–1123. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- BBS/B/03858/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- P41 RR011823/RR/NCRR NIH HHS/United States
- R21 AI072615/AI/NIAID NIH HHS/United States
- BBS/B/03742/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- R21 AI072615-01/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
