

Review
doi: 10.1111/j.1399-3089.2009.00534.x.
Current status of xenotransplantation and prospects for clinical application
Affiliations
- PMID: 19796067
- PMCID: PMC2866107
- DOI: 10.1111/j.1399-3089.2009.00534.x
Item in Clipboard
Review
Current status of xenotransplantation and prospects for clinical application
Xenotransplantation.
2009 Sep-Oct.
Abstract
Xenotransplantation is one promising approach to bridge the gap between available human cells, tissues, and organs and the needs of patients with diabetes or end-stage organ failure. Based on recent progress using genetically modified source pigs, improving results with conventional and experimental immunosuppression, and expanded understanding of residual physiologic hurdles, xenotransplantation appears likely to be evaluated in clinical trials in the near future for some select applications. This review offers a comprehensive overview of known mechanisms of xenograft injury, a contemporary assessment of preclinical progress and residual barriers, and our opinions regarding where breakthroughs are likely to occur.
Figures

According to the conventional model, hyperacute rejection of pig organs in humans is triggered by binding of preformed human “natural” antibody, which is predominantly directed against Galα1-3Gal sugars that decorate many molecules on the surface of pig cells. Panel A : Complement is activated by bound antibody, triggering retraction and lysis of endothelial cells. Loss of endothelial barrier function contributes to leakage of blood into the interstitium. Blood clotting within vessels is activated by injured endothelium and exposed basement membrane, leading to downstream tissue ischemia and necrosis. Panel B: When the galactosyl transferase gene is disabled by gene knockout, pig cells lack the Galα1-3Gal target antigen. If present in sufficient quantity, antibody against other pig antigens that is either preformed or induced after transplant can trigger complement-mediated injury and associated coagulation pathway activation. Panel C: Human proteins that regulate (abort) complement activation (complement pathway regulatory proteins, CPRPs) disable human complement more efficiently than do their pig analogues. Organs from pigs genetically engineered to express human CPRPs are protected from hyperacute rejection, although this protection can be overcome by high-titer antibody, or if endothelial injury occurs due to other mechanisms (ischemia-perfusion injury or coagulation pathway activation, for example). GalT-KO, human CPRP, and perhaps additional gene modifications may be necessary to yield clinically useful, reliable protection of pig organs in man.

According to the conventional model, hyperacute rejection of pig organs in humans is triggered by binding of preformed human “natural” antibody, which is predominantly directed against Galα1-3Gal sugars that decorate many molecules on the surface of pig cells. Panel A : Complement is activated by bound antibody, triggering retraction and lysis of endothelial cells. Loss of endothelial barrier function contributes to leakage of blood into the interstitium. Blood clotting within vessels is activated by injured endothelium and exposed basement membrane, leading to downstream tissue ischemia and necrosis. Panel B: When the galactosyl transferase gene is disabled by gene knockout, pig cells lack the Galα1-3Gal target antigen. If present in sufficient quantity, antibody against other pig antigens that is either preformed or induced after transplant can trigger complement-mediated injury and associated coagulation pathway activation. Panel C: Human proteins that regulate (abort) complement activation (complement pathway regulatory proteins, CPRPs) disable human complement more efficiently than do their pig analogues. Organs from pigs genetically engineered to express human CPRPs are protected from hyperacute rejection, although this protection can be overcome by high-titer antibody, or if endothelial injury occurs due to other mechanisms (ischemia-perfusion injury or coagulation pathway activation, for example). GalT-KO, human CPRP, and perhaps additional gene modifications may be necessary to yield clinically useful, reliable protection of pig organs in man.
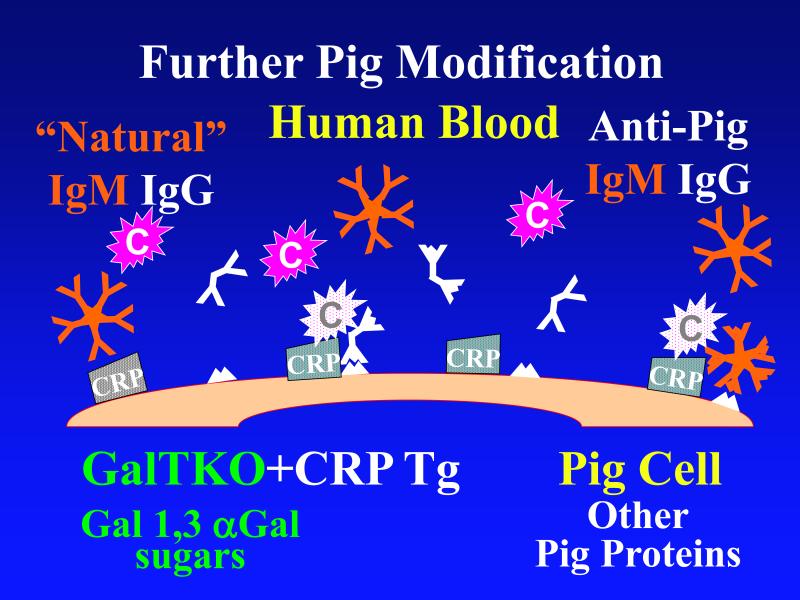
According to the conventional model, hyperacute rejection of pig organs in humans is triggered by binding of preformed human “natural” antibody, which is predominantly directed against Galα1-3Gal sugars that decorate many molecules on the surface of pig cells. Panel A : Complement is activated by bound antibody, triggering retraction and lysis of endothelial cells. Loss of endothelial barrier function contributes to leakage of blood into the interstitium. Blood clotting within vessels is activated by injured endothelium and exposed basement membrane, leading to downstream tissue ischemia and necrosis. Panel B: When the galactosyl transferase gene is disabled by gene knockout, pig cells lack the Galα1-3Gal target antigen. If present in sufficient quantity, antibody against other pig antigens that is either preformed or induced after transplant can trigger complement-mediated injury and associated coagulation pathway activation. Panel C: Human proteins that regulate (abort) complement activation (complement pathway regulatory proteins, CPRPs) disable human complement more efficiently than do their pig analogues. Organs from pigs genetically engineered to express human CPRPs are protected from hyperacute rejection, although this protection can be overcome by high-titer antibody, or if endothelial injury occurs due to other mechanisms (ischemia-perfusion injury or coagulation pathway activation, for example). GalT-KO, human CPRP, and perhaps additional gene modifications may be necessary to yield clinically useful, reliable protection of pig organs in man.
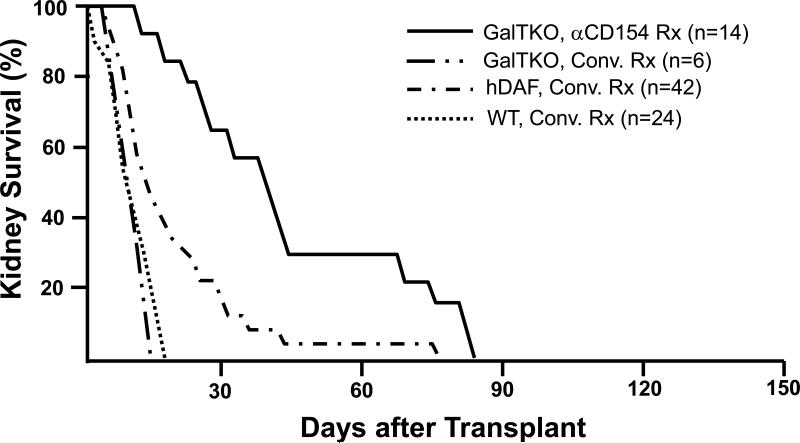
Survival of life-supporting kidneys from wild-type (WT, n=24), hDAF transgenic (n=42), or GalT-KO pigs (n=20) in immunosuppressed baboons. With intensive conventional immunosuppression, with or without additional complement depletion, hDAF kidney recipients succumbed to infectious complications or developed DXR at a median of 15 to 25 days, significantly later than do similarly treated recipients of wild-type organs (~6-10 days). The incidence of early graft failure (<3 days) was reduced in recipients of GalT-KO kidneys. Six GalT-KO kidneys in baboons treated with conventional immunosuppression exhibited DXR associated with induction of anti-pig antibodies, demonstrating that the regimens tested in this experience were subtherapeutic. Using an anti-CD154 antibody, mycophenolate mofetil, T-cell depletion, and vascularized donor thymus tissue, 8 of 14 recipient baboons in the GalT-KO kidney group died with functioning grafts (serum creatinine <2.0mg/dL) from treatment complications. (Results in 3 recipients of hDAF kidneys treated with anti-CD154 antibody, which survived for 28, 29 and 29 days, are omitted from the figure due to the relatively small experience.) Data for conventional immunosuppression is abstracted and aggregated from all peer-reviewed pig-to-baboon kidney transplant studies published since 1998. Pig phenotype (GalT-KO, hDAF, or WT) and treatment regimen (anti-CD154-based or conventional (Conv.) treatment (Rx)) are indicated. Similar results were achieved with conventional immunosuppression in over 250 cynomolgus recipients of hDAF kidneys that used comparable approaches.
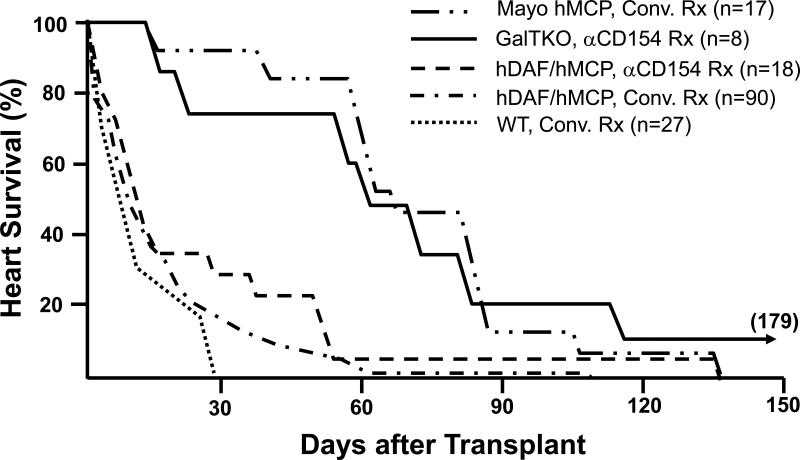
Survival in immunosuppressed baboons of heterotopic hearts from wild-type (WT) pigs (n=26), pigs transgenic for human decay-accelerating factor (hDAF), human membrane cofactor protein (hMCP), and/or human CD59 (n=125), and GalT-KO pigs (n=8). In conjunction with neutralization of anti-Gal antibodies, conventional or anti-CD154-based immunosuppression (using a monoclonal antibody that selectively blocks the CD154-CD40 “costimulation” pathway important to T- and B-cell activation) was associated with median survival of hDAF or hMCP transgenic hearts approaching one month in some series, and for over 2 months in the Mayo Clinic's recent experience. Infectious or treatment-associated complications caused the death of over 50% of recipients treated with conventional immunosuppression, but were unusual with anti-CD154-based treatment. Lymphocyte and antibody responses against pig antigens were generally undetectable during either anti-CD154 or intensive conventional therapy. Grafts failed with DXR histology in association with both conventional and CD154-based treatment regimens. Eight GalT-KO hearts in baboons treated with anti-CD154 antibody, mycophenlate mofetil, and perioperative T-cell depletion, did not exhibit early graft failure (<3 days), and median survival exceeded two months. Typical DXR was rare in GalT-KO hearts, but thrombotic microangiopathy, which was also observed in long-surviving hDAF and hMCP hearts, was prominent in association with GalT-KO cardiac xenograft failure. Data are abstracted from all peer-reviewed pig-to-baboon heart transplant studies published since 1988, aggregating data from studies that used similar therapeutic approaches. Pig phenotype (GalT-KO, hDAF, hMCP, or WT) and treatment regimen (anti-CD154-based or conventional (Conv.) treatment (Rx)) are indicated. Similar results were achieved with conventional immunosuppression in over 65 cynomolgus recipients of hDAF hearts.
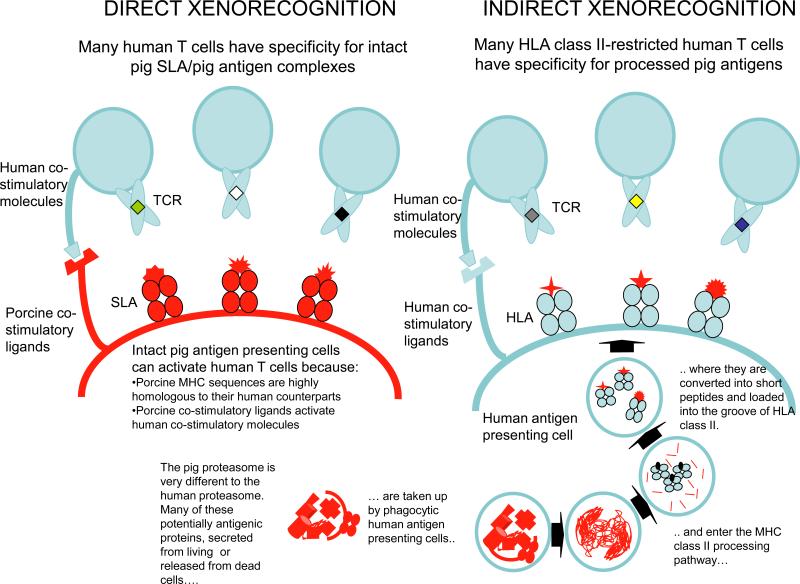
Available routes for presentation of pig antigens to primate T-cells (Editor's note: Legend is incorporate into the figure.)
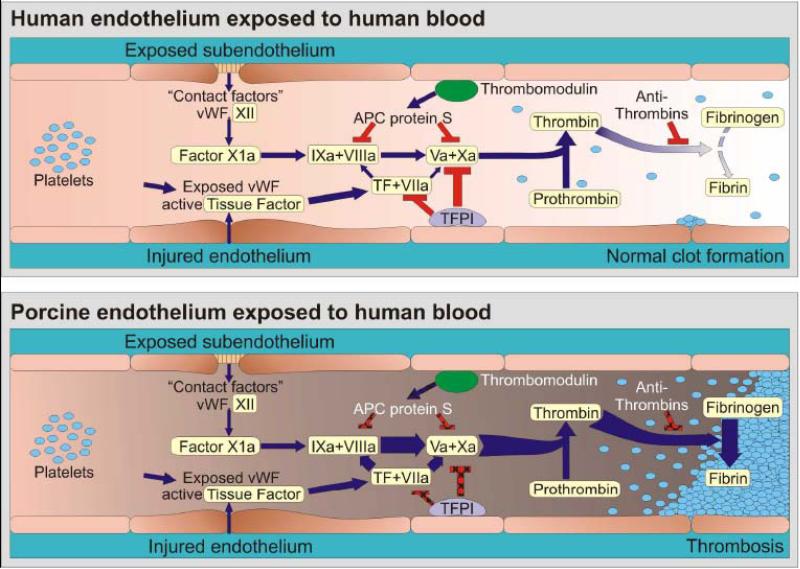
Coagulation is occurring continuously within the blood stream, but is normally restrained by a network of inhibitory pathways involving endothelial proteins such as thrombomodulin and tissue factor pathway inhibitor (TFPI) (Panel A ). Increased coagulation is normally initiated when endothelium retracts or becomes “activated” by injury, in part because von Willebrand factor (vWF) is expressed and tissue factor (TF) is liberated into the circulation. The coagulation cascade then becomes amplified by the factors shown (VIIa/TF complex, IXa and Xa) which in turn activate thrombin. Thrombin amplifies the clotting cascade by a) activating XIa (not shown), b) activating platelets, c) cleaving fibrinogen into fibrin monomers that form the primary clot matrix, and d) activating factor XIIIa (not shown), which cross-links fibrin monomers into an insoluble clot. TFPI and thrombomodulin normally inhibit coagulation on healthy endothelium, while soluble antithrombins inhibit thrombin by forming a complex with its active site. Porcine EC activation – whether by xenoantibodies, complement, or other factors – results in loss of natural anticoagulant proteins (TFPI, thrombomodulin) and acquisition of a procoagulant phenotype (Panel B ). In addition functional incompatibilities in the coagulation system between pigs and humans cause both inappropriate or accelerated thrombin formation and inefficient restraint of clot activation. Our current hypothesis is that xenografts succumb to an otherwise insignificant humoral or cellular immune response which amplifies endothelial injury and intravascular thrombosis, and becomes manifest as TM. Blue arrows designate cascade amplification steps, while red lines identify loci of inhibition. The relative intensity of clot formation, the net product of coagulation pathway enzyme effects, is symbolized by arrow weight at the thrombin and fibrin steps. Pathways where pig endothelial proteins inefficiently dampen coagulation are indicated with hatchmarked red lines in Panel B. For simplicity, only the activated clotting factor intermediaries and key points at which regulation occurs are shown.
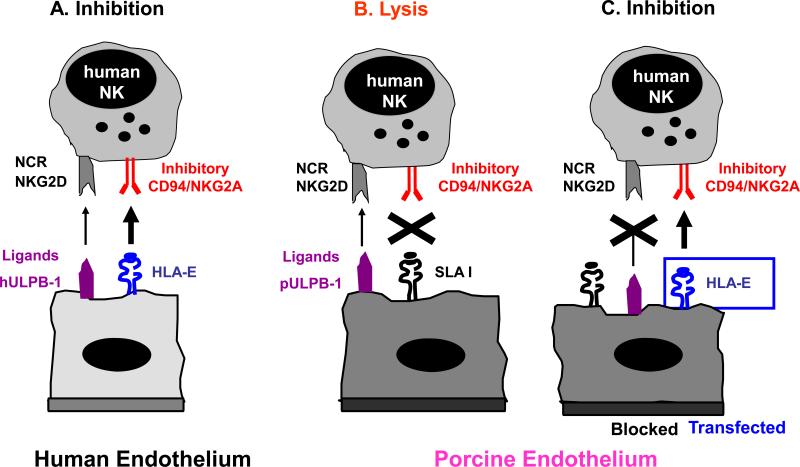
Molecular interactions between human NK cells and pig endothelial cells NK cells are tightly regulated through signals mediated by inhibiting and activating receptors expressed on their cell-surface. Many of the inhibitory NK receptors recognize MHC class I molecules and therefore allow NK cells to discriminate between self and non-self. Activating receptors on NK cells include natural cytotoxicity receptors (NCR) and the C-type lectin receptor NKG2D. (A) Human endothelium is normally protected from NK cytotoxicity by dominant inhibitory signals provided by HLA, e.g. recognition of HLA-E by CD94/NKG2A. (B) In contrast, porcine endothelium is susceptible to human NK cell-mediated lysis likely due to the inability of swine leukocyte antigen (SLA) MHC class I molecules to signal through human NK inhibitory receptors. Human NK cytotoxicity against porcine EC is mediated primarily through NKG2D-pULBP-1 interactions and the NCR NKp44. (C) Transgenic expression of HLA class I molecules on porcine endothelium resulted in partial protection whereas additional blocking of activating NK receptors and their porcine ligands provides complete protection from human NK cytotoxicity.
References
-
- Cooper DK, Dorling A, Pierson RN, III, et al. Alpha1,3-galactosyltransferase gene knockout pigs for xenotransplantation: where do we go from here? Transplantation. 2007;84(1):1–7. - PubMed
-
- Allan JS. The risk of using baboons as transplant donors: exogenous and endogenous viruses. Ann N Y Acad Sci. 1998;862:87–99. - PubMed
-
- Goodall J. Ethical concerns in the use of animals as donors. In: Hardy MA, editor. Xenograft 25. Excerpta Medica International Congress Series, Elsevier Science; Amsterdam, Netherlands: 1989. pp. 335–349.
-
- Byrne GW, McCurry KR, Martin MJ, McClellan SM, Platt JL, Logan JS. Transgenic pigs expressing human CD59 and decay-accelerating factor produce an intrinsic barrier to complement-mediated damage. Transplantation. 1997;63(1):149–55. - PubMed
-
- Langford GA, Yannoutsos N, Cozzi E, et al. Production of pigs transgenic for human decay accelerating factor. Transplant Proc. 1994;26(3):1400–1. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- UO1 AI068642/AI/NIAID NIH HHS/United States
- U01 AI 066719-01/AI/NIAID NIH HHS/United States
- P01-AI45897/AI/NIAID NIH HHS/United States
- U19 AI067151/AI/NIAID NIH HHS/United States
- P01 AI045897/AI/NIAID NIH HHS/United States
- U01 AI066719/AI/NIAID NIH HHS/United States
- U01 AI068642/AI/NIAID NIH HHS/United States
- U01 AI066335/AI/NIAID NIH HHS/United States
- UO1 AI 066335-01/AI/NIAID NIH HHS/United States
- R01 DK066160/DK/NIDDK NIH HHS/United States
- R21 A1074844/PHS HHS/United States
- G0801965/MRC_/Medical Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
