

Pulmonary alveolar proteinosis, a primary immunodeficiency of impaired GM-CSF stimulation of macrophages
- PMID: 19796925
- PMCID: PMC2779868
- DOI: 10.1016/j.coi.2009.09.004
Pulmonary alveolar proteinosis, a primary immunodeficiency of impaired GM-CSF stimulation of macrophages
Abstract
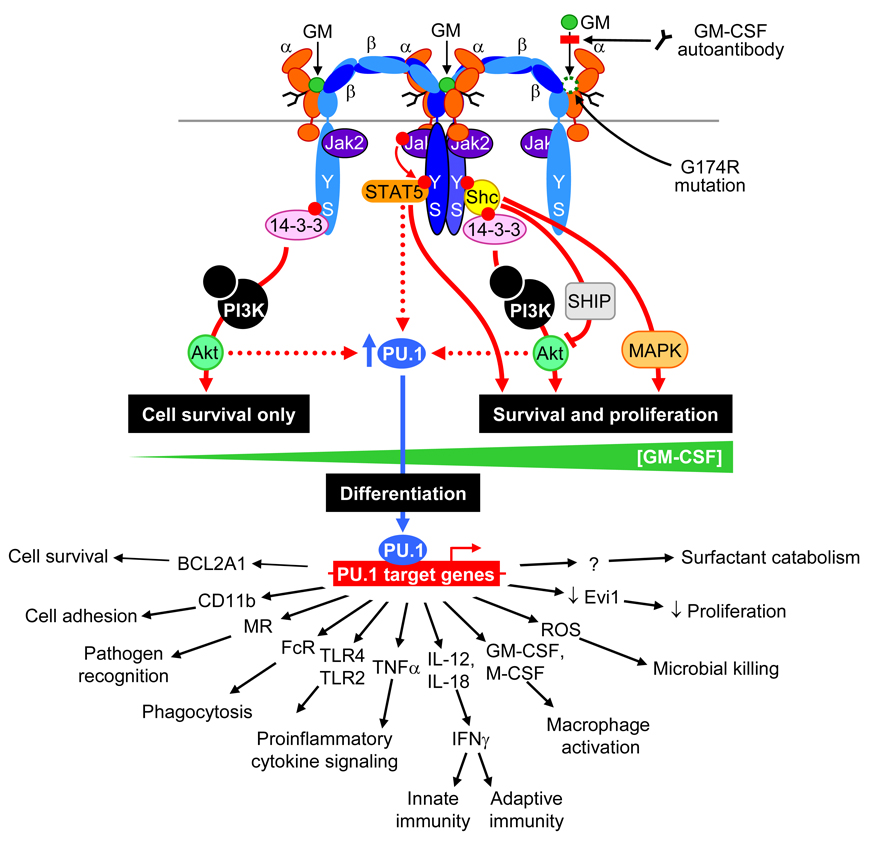
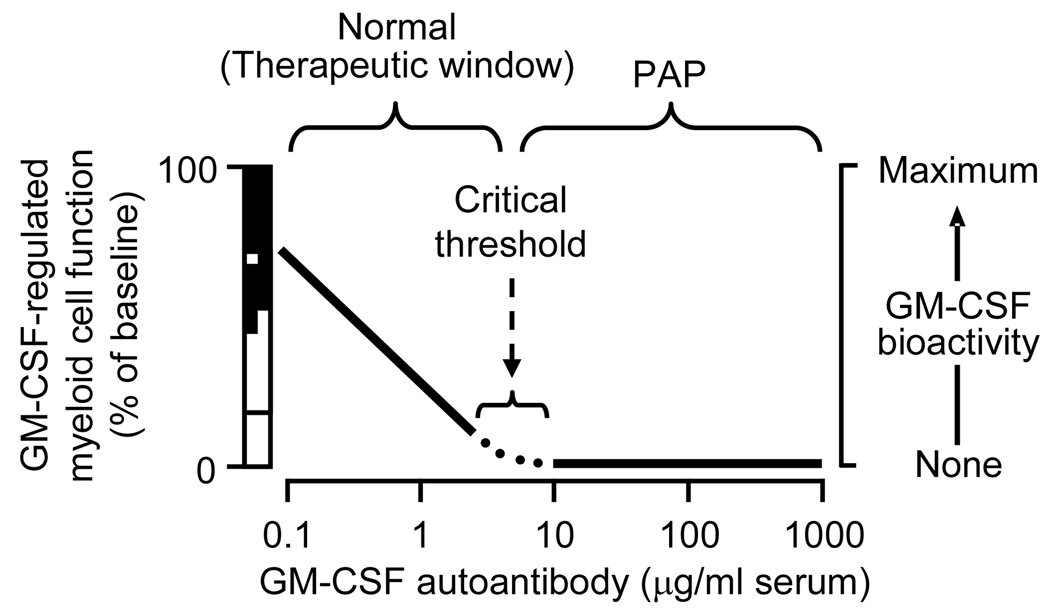
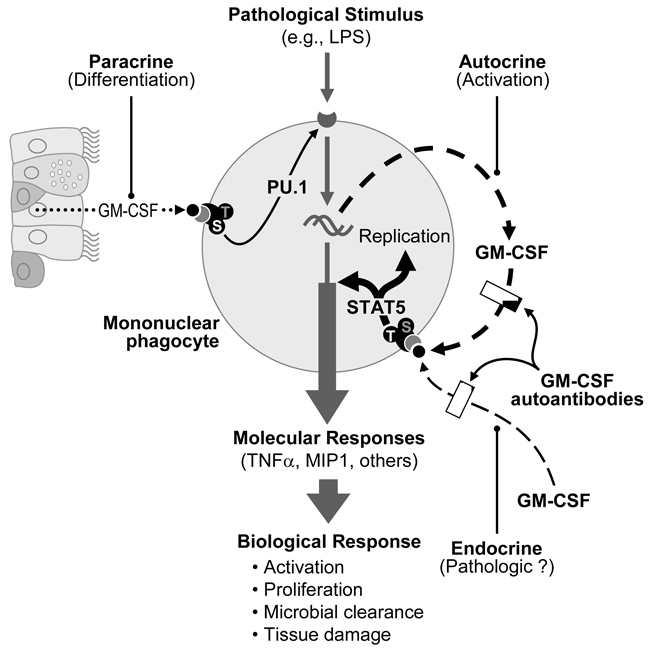
Pulmonary alveolar proteinosis (PAP) is a rare syndrome characterized by accumulation of pulmonary surfactant, respiratory insufficiency, and increased infections. It occurs in various clinical settings that disrupt surfactant catabolism in alveolar macrophages, including a relatively more common autoimmune disease caused by GM-CSF autoantibodies and a rare congenital disease caused by CSF2RA mutations. Recent results demonstrate that GM-CSF is crucial for alveolar macrophage terminal differentiation and immune functions, pulmonary surfactant homeostasis, and lung host defense. GM-CSF is also required to determine the basal functional capacity of circulating neutrophils, including adhesion, phagocytosis, and microbial killing. PAP research has illuminated the crucial role of GM-CSF in innate immunity and led to novel therapy for PAP and the potential use of anti-GM-CSF therapy in other common disorders.
Figures
References
-
-
Hercus TR, Thomas D, Guthridge MA, Ekert PG, King-Scott J, Parker MW, Lopez AF. The GM-CSF receptor: linking its structure to cell signaling and its role in disease. Blood. 2009 This paper presents a new model for signaling by the GM-CSF receptor, based on the recently elucidated dodecameric structure of GM-CSF bound to complexes of the GM-CSF receptor α and β chains, and Jak2. This new structure provides important new insights into the pleiotropic effects of GM-CSF derived from an understanding of the receptor’s structure.
-
-
- Trapnell BC, Whitsett JA. GM-CSF regulates pulmonary surfactant homeostasis and alveolar macrophage-mediated innate host defense. Annu Rev Physiol. 2002;64:775–802. - PubMed
-
- Presneill JJ, Nakata K, Inoue Y, Seymour JF. Pulmonary alveolar proteinosis. Clin Chest Med. 2004;25:593–613. viii. - PubMed
-
- Trapnell BC, Whitsett JA, Nakata K. Pulmonary Alveolar Proteinosis. N Engl J Med. 2003;349:2527–2539. - PubMed
-
-
Seymour JF, Presneill JJ. Pulmonary alveolar proteinosis: progress in the first 44 years. Am J Respir Crit Care Med. 2002;166:215–235. This study provides a comprehensive metaanalysis summarizing most or all of the cases of PAP reported in the medical literature from the initial description of the syndrome in 1958 through 1997.
-
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
