

Toll-like receptor signaling in airborne Burkholderia thailandensis infection
- PMID: 19797072
- PMCID: PMC2786489
- DOI: 10.1128/IAI.00618-09
Toll-like receptor signaling in airborne Burkholderia thailandensis infection
Abstract
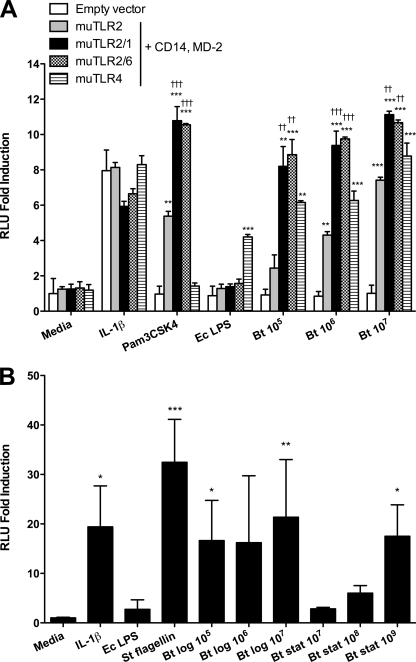
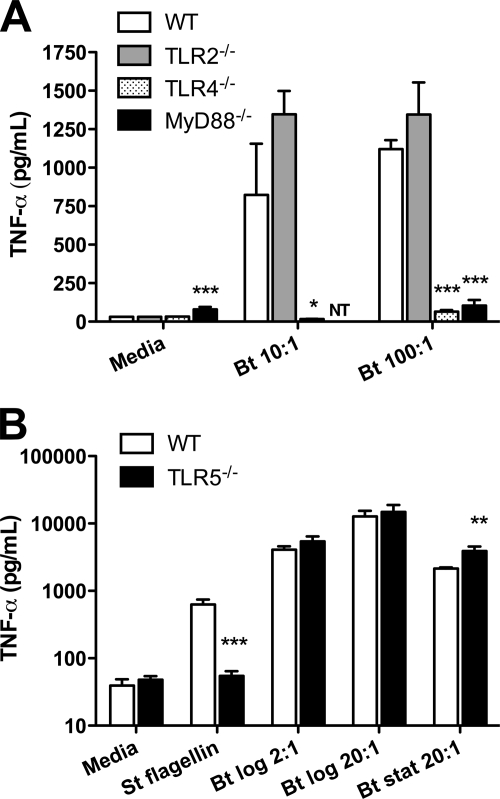
Melioidosis is a tropical disease endemic in southeast Asia and northern Australia caused by the gram-negative soil saprophyte Burkholderia pseudomallei. Although infection is often systemic, the lung is frequently involved. B. thailandensis is a closely related organism that at high doses causes lethal pneumonia in mice. We examined the role of Toll-like receptors (TLRs), essential components of innate immunity, in vitro and in vivo during murine B. thailandensis pneumonia. TLR2, TLR4, and TLR5 mediate NF-kappaB activation by B. thailandensis in transfected HEK293 or CHO cells. In macrophages, TLR4 and the adaptor molecule MyD88, but not TLR2 or TLR5, are required for tumor necrosis factor alpha production induced by B. thailandensis. In low-dose airborne infection, TLR4 is needed for early, but not late, bacterial containment, and MyD88 is essential for control of infection and host survival. TLR2 and TLR5 are not necessary to contain low-dose infection. In high-dose airborne infection, TLR2 deficiency confers a slight survival advantage. Lung and systemic inflammatory responses are induced by low-dose inhaled B. thailandensis independently of individual TLRs or MyD88. These findings suggest that redundancy in TLR signaling or other MyD88-dependent pathways may be important in pneumonic B. thailandensis infection but that MyD88-independent mechanisms of inflammation are also activated. TLR signaling in B. thailandensis infection is substantially comparable to signaling induced by virulent B. pseudomallei. These studies provide additional insights into the host-pathogen interaction in pneumonic Burkholderia infection.
Figures
Similar articles
-
Activation of Toll-like receptors by Burkholderia pseudomallei.BMC Immunol. 2008 Aug 8;9:46. doi: 10.1186/1471-2172-9-46. BMC Immunol. 2008. PMID: 18691413 Free PMC article.
-
Differential susceptibility of inbred mouse strains to Burkholderia thailandensis aerosol infection.Microb Pathog. 2010 Jan;48(1):9-17. doi: 10.1016/j.micpath.2009.10.004. Epub 2009 Oct 21. Microb Pathog. 2010. PMID: 19853031 Free PMC article.
-
Burkholderia pseudomallei-derived miR-3473 enhances NF-κB via targeting TRAF3 and is associated with different inflammatory responses compared to Burkholderia thailandensis in murine macrophages.BMC Microbiol. 2016 Nov 28;16(1):283. doi: 10.1186/s12866-016-0901-6. BMC Microbiol. 2016. PMID: 27894256 Free PMC article.
-
Role of Canonical and Non-canonical Inflammasomes During Burkholderia Infection.Curr Top Microbiol Immunol. 2016;397:199-214. doi: 10.1007/978-3-319-41171-2_10. Curr Top Microbiol Immunol. 2016. PMID: 27460811 Review.
-
Membrane TLR signaling mechanisms in the gastrointestinal tract during sepsis.Neurogastroenterol Motil. 2010 Mar;22(3):232-45. doi: 10.1111/j.1365-2982.2009.01464.x. Neurogastroenterol Motil. 2010. PMID: 20377787 Free PMC article. Review.
Cited by
-
Survey of innate immune responses to Burkholderia pseudomallei in human blood identifies a central role for lipopolysaccharide.PLoS One. 2013 Nov 26;8(11):e81617. doi: 10.1371/journal.pone.0081617. eCollection 2013. PLoS One. 2013. PMID: 24303060 Free PMC article.
-
Lactoferrin is a dynamic protein in human melioidosis and is a TLR4-dependent driver of TNF-α release in Burkholderia thailandensis infection in vitro.PLoS Negl Trop Dis. 2020 Aug 7;14(8):e0008495. doi: 10.1371/journal.pntd.0008495. eCollection 2020 Aug. PLoS Negl Trop Dis. 2020. PMID: 32764765 Free PMC article.
-
Toll-like receptor 4 region genetic variants are associated with susceptibility to melioidosis.Genes Immun. 2012 Jan;13(1):38-46. doi: 10.1038/gene.2011.49. Epub 2011 Jul 21. Genes Immun. 2012. PMID: 21776015 Free PMC article.
-
Impaired TLR5 functionality is associated with survival in melioidosis.J Immunol. 2013 Apr 1;190(7):3373-9. doi: 10.4049/jimmunol.1202974. Epub 2013 Feb 27. J Immunol. 2013. PMID: 23447684 Free PMC article.
-
Sequence-defined transposon mutant library of Burkholderia thailandensis.mBio. 2013 Nov 5;4(6):e00604-13. doi: 10.1128/mBio.00604-13. mBio. 2013. PMID: 24194535 Free PMC article.
References
-
- Bhan, U., N. W. Lukacs, J. J. Osterholzer, M. W. Newstead, X. Zeng, T. A. Moore, T. R. McMillan, A. M. Krieg, S. Akira, and T. J. Standiford. 2007. TLR9 is required for protective innate immunity in gram-negative bacterial pneumonia: role of dendritic cells. J. Immunol. 179:3937-3946. - PubMed
-
- Bossi, P., A. Tegnell, A. Baka, F. Van Loock, J. Hendriks, A. Werner, H. Maidhof, and G. Gouvras. 2004. Bichat guidelines for the clinical management of glanders and melioidosis and bioterrorism-related glanders and melioidosis. Euro Surveill. 9:E17-E18. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
