

Review
doi: 10.1083/jcb.200907014.
Coordination of membrane events during autophagy by multiple class III PI3-kinase complexes
Affiliations
- PMID: 19797076
- PMCID: PMC2753151
- DOI: 10.1083/jcb.200907014
Item in Clipboard
Review
Coordination of membrane events during autophagy by multiple class III PI3-kinase complexes
J Cell Biol.
.
Abstract
Autophagy or "self-eating" is a highly conserved pathway that enables cells to degrade pieces of themselves in autolysosomes to enable their survival in times of stress, including nutrient deprivation. The formation of these degradative compartments requires cytosolic proteins, some of which are autophagy specific, as well as intracellular organelles, such as the ER and Golgi, and the endosome-lysosome system. Here we discuss the cross talk between autophagy and intracellular compartments, highlighting recent exciting data about the role and regulation of the Vps34 class III phosphatidylinositol (PI) 3-kinase in autophagy.
Figures
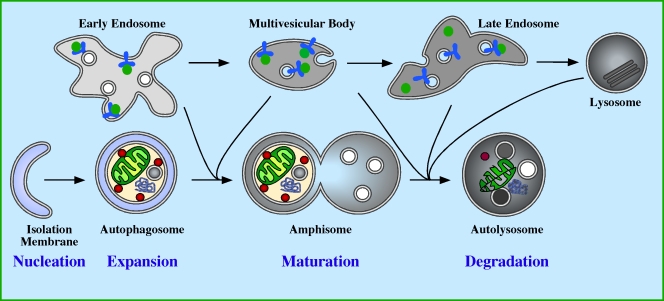
Autophagy in mammalian cells. The autophagic pathway comprises four stages: nucleation, expansion, maturation, and degradation. Nucleation occurs in response to a signal transmitted downstream of a cellular stress, causing the isolation membrane (IM) to start to grow. The IM is also known as the phagophore (Seglen and Bohley, 1992). Expansion of the IM occurs, enabling it to grow in all dimensions sequestering cytosolic proteins, organelles, and aggregated proteins. Expansion is complete when the membrane closes to form the double membrane autophagosome vesicle. After closure of the autophagosome, maturation begins by fusion with endocytic compartments, including early endosomes, multivesicular bodies, late endosomes, and lysosomes. The step-wise, or individual fusion events of the autophagosome with the endocytic compartment create an amphisome that has both autophagosomal and endosomal content. During maturation the lumen of the amphisome acidifies, and the membrane acquires LAMPs (lysosmomal membrane proteins), hydrolytic enzymes, and lipases. This leads to the formation of an autolysosome, within which the sequestered content is degraded and recycled back to the cytosol.
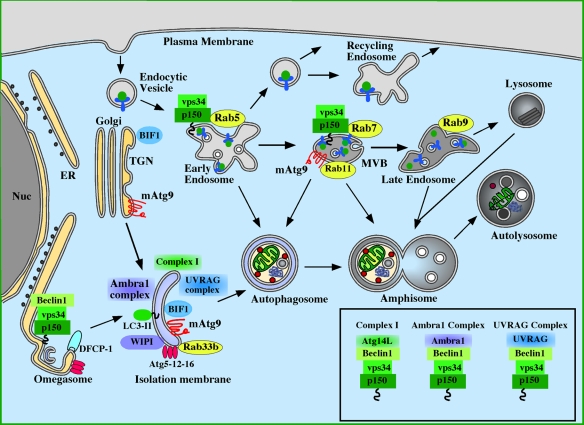
Molecules involved in the induction of autophagy: Atg proteins, Rab proteins, and the class III PI3 kinase. The source of the isolation membrane is unknown, but has been proposed to be derived from the endoplasmic reticulum (ER) or trans-Golgi network (TGN). The omegasome is continuous with the ER and is also proposed to contribute to the IM formation. Class III PI3K complex I (p150–Vps34–Beclin1–Atg14L) is localized to the IM and is required for autophagy (Itakura et al., 2008; Sun et al., 2008; Matsunaga et al., 2009; Zhong et al., 2009). A subset of this complex (p150-Vps34) is required for endocytosis and is found on early endosomes and multivesicular bodies (MVBs), and interacts with Rab5 and Rab7. A Beclin1-containing subcomplex is required for function of the omegasome and the recruitment of DFCP-1 (Axe et al., 2008). Recent data also point to two additional p150–Vps34–Beclin1 complexes, Ambra1 and UVRAG complexes, being required for autophagy. Ambra1 associates directly with Beclin1 and is required for induction of autophagy (Fimia et al., 2007). UVRAG likewise associates directly with Beclin1, and is required for autophagy (Liang et al., 2008). BIF-1, previously shown to be required for fission of Golgi carriers (Yang et al., 2006), associates with UVRAG and has been shown to be required for autophagy (Takahashi et al., 2007). The activity of the PI3K Vps34 is enhanced by interaction with Rab5, Rab7, and Beclin1-associated UVRAG together with BIF-1, but it is not known how Ambra1 association with Beclin1 affects the activity of the PI3K. The composition of these complexes is shown in the box.
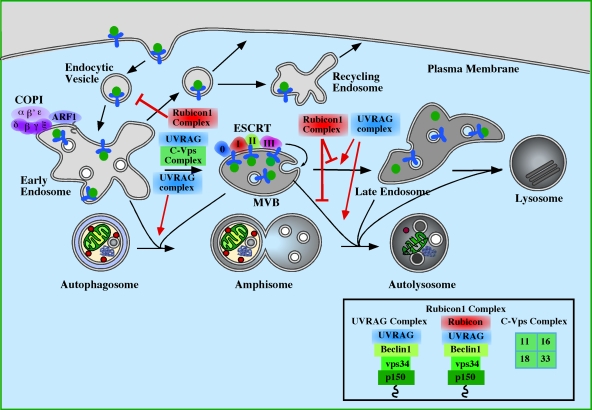
The role of endosomes and Class III PI3-kinase complexes in autophagosome maturation. Maturation of autophagosomes has been shown to require functional early endosomes, multivesicular bodies (MVBs) and late endosomes. COPI is important for early steps in endocytosis and for autophagosome maturation (Razi et al., 2009). Similarly, the ESCRT complexes participate in autophagosome maturation and later steps of endocytosis (Raiborg and Stenmark, 2009). Loss of either COPI or ESCRT results in the accumulation of autophagosomes and amphisomes. Loss of the Class III PI3K subcomplex p150-Vps34 (see Fig. 2) would also be predicted to have a similar inhibition of maturation as it is required to maintain endosomal function on Rab5- and Rab7-positive endosomes. Of the Class III PI3K complexes, the UVRAG complex, described in Fig. 2 and the Rubicon complex (p150–Vps34–Beclin1–UVRAG–Rubicon) regulate autophagosome maturation (Matsunaga et al., 2009; Zhong et al., 2009). UVRAG, required for endosome function, positively regulates maturation and autophagy, while the association of Rubicon to UVRAG causes an inhibition of autophagic maturation. Note: Rubicon complex also reduces the activity of the Class III PI3K. The composition of the complexes is shown in the box. UVRAG can also associate with the core class C–Vps complex, containing Vps11, 16, 18, and 33, independently of Beclin1, and this interaction accelerates autophagy (Liang et al., 2008), perhaps though stimulation of Rab7 activity.
References
-
- Audhya A., Desai A., Oegema K. 2007. A role for Rab5 in structuring the endoplasmic reticulum.J. Cell Biol. 178:43–56 doi:10.1083/jcb.200701139 - DOI - PMC - PubMed
-
- Axe E.L., Walker S.A., Manifava M., Chandra P., Roderick H.L., Habermann A., Griffiths G., Ktistakis N.T. 2008. Autophagosome formation from membrane compartments enriched in phosphatidylinositol 3-phosphate and dynamically connected to the endoplasmic reticulum.J. Cell Biol. 182:685–701 doi:10.1083/jcb.200803137 - DOI - PMC - PubMed
-
- Babst M., Wendland B., Estepa E.J., Emr S.D. 1998. The Vps4p AAA ATPase regulates membrane association of a Vps protein complex required for normal endosome function.EMBO J. 17:2982–2993 doi:10.1093/emboj/17.11.2982 - DOI - PMC - PubMed
-
- Backer J.M. 2008. The regulation and function of Class III PI3Ks: novel roles for Vps34.Biochem. J. 410:1–17 doi:10.1042/BJ20071427 - DOI - PubMed
-
- Burda P., Padilla S.M., Sarkar S., Emr S.D. 2002. Retromer function in endosome-to-Golgi retrograde transport is regulated by the yeast Vps34 PtdIns 3-kinase.J. Cell Sci. 115:3889–3900 doi:10.1242/jcs.00090 - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
