

Analysis of lipid transfer activity between model nascent HDL particles and plasma lipoproteins: implications for current concepts of nascent HDL maturation and genesis
- PMID: 19797257
- PMCID: PMC2842159
- DOI: 10.1194/jlr.M001875
Analysis of lipid transfer activity between model nascent HDL particles and plasma lipoproteins: implications for current concepts of nascent HDL maturation and genesis
Abstract
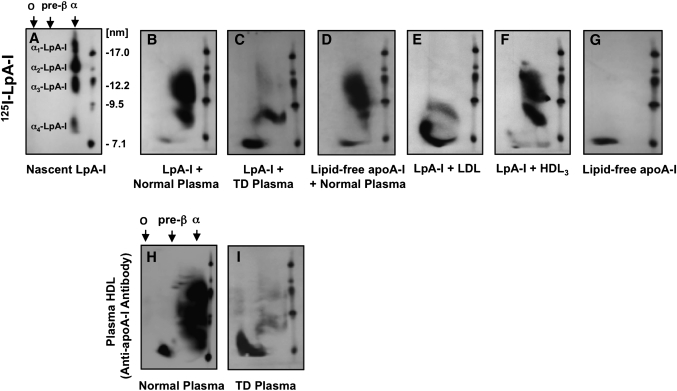
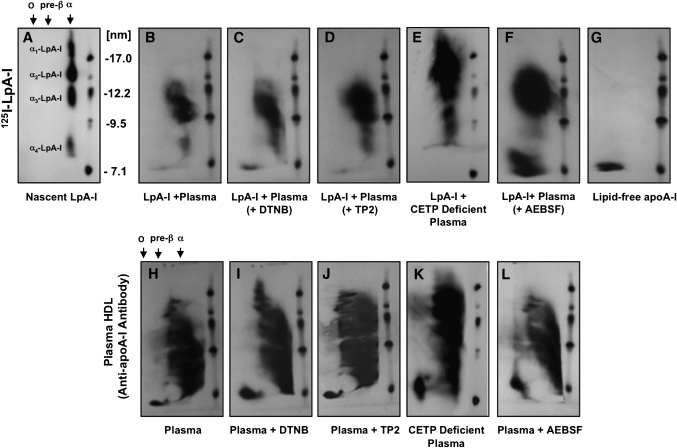
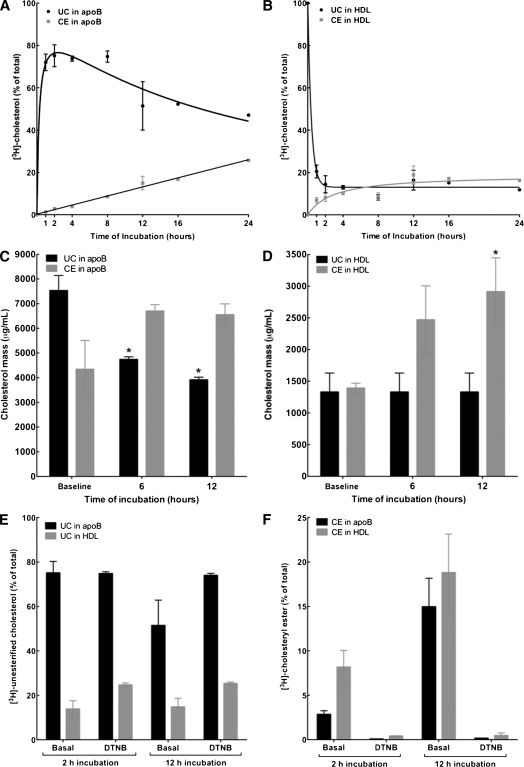
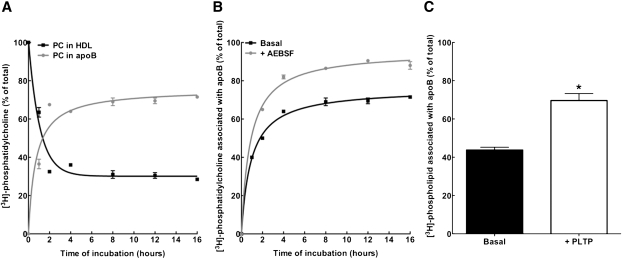
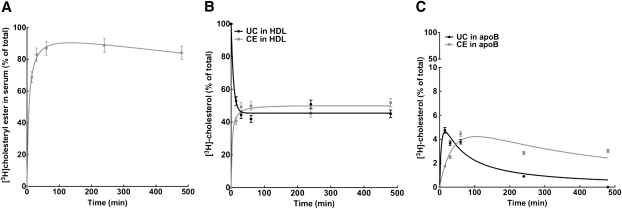
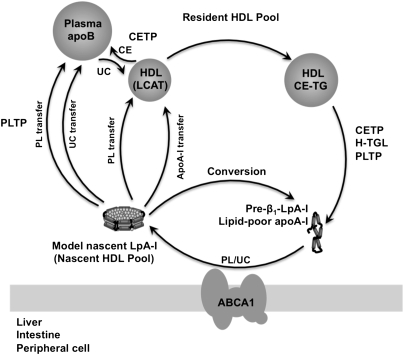
The specifics of nascent HDL remodeling within the plasma compartment remain poorly understood. We developed an in vitro assay to monitor the lipid transfer between model nascent HDL (LpA-I) and plasma lipoproteins. Incubation of alpha-(125)I-LpA-I with plasma resulted in association of LpA-I with existing plasma HDL, whereas incubation with TD plasma or LDL resulted in conversion of alpha-(125)I-LpA-I to prebeta-HDL. To further investigate the dynamics of lipid transfer, nascent LpA-I were labeled with cell-derived [(3 )H]cholesterol (UC) or [(3)H]phosphatidylcholine (PC) and incubated with plasma at 37 degrees C. The majority of UC and PC were rapidly transferred to apolipoprotein B (apoB). Subsequently, UC was redistributed to HDL for esterification before being returned to apoB. The presence of a phospholipid transfer protein (PLTP) stimulator or purified PLTP promoted PC transfer to apoB. Conversely, PC transfer was abolished in plasma from PLTP(-/-) mice. Injection of (125)I-LpA-I into rabbits resulted in a rapid size redistribution of (125)I-LpA-I. The majority of [(3)H]UC from labeled r(HDL) was esterified in vivo within HDL, whereas a minority was found in LDL. These data suggest that apoB plays a major role in nascent HDL remodeling by accepting their lipids and donating UC to the LCAT reaction. The finding that nascent particles were depleted of their lipids and remodeled in the presence of plasma lipoproteins raises questions about their stability and subsequent interaction with LCAT.
Figures



Similar articles
-
Reverse cholesterol transport in plasma of patients with different forms of familial HDL deficiency.Arterioscler Thromb Vasc Biol. 1995 May;15(5):691-703. doi: 10.1161/01.atv.15.5.691. Arterioscler Thromb Vasc Biol. 1995. PMID: 7749883
-
A natural apolipoprotein A-I variant, apoA-I (L141R)Pisa, interferes with the formation of alpha-high density lipoproteins (HDL) but not with the formation of pre beta 1-HDL and influences efflux of cholesterol into plasma.J Lipid Res. 1997 Jun;38(6):1242-53. J Lipid Res. 1997. PMID: 9215551
-
Phospholipid transfer protein mediated conversion of high density lipoproteins generates pre beta 1-HDL.Biochim Biophys Acta. 1996 Jun 11;1301(3):255-62. doi: 10.1016/0005-2760(96)00050-1. Biochim Biophys Acta. 1996. PMID: 8664337
-
The origin and metabolism of a nascent pre-β high density lipoprotein involved in cellular cholesterol efflux.Acta Biochim Pol. 2011;58(3):275-85. Epub 2011 Jul 12. Acta Biochim Pol. 2011. PMID: 21750785 Review.
-
High density lipoproteins and reverse cholesterol transport: lessons from mutations.Atherosclerosis. 1998 Apr;137 Suppl:S7-11. doi: 10.1016/s0021-9150(97)00311-0. Atherosclerosis. 1998. PMID: 9694535 Review.
Cited by
-
PLTP activity inversely correlates with CAAD: effects of PON1 enzyme activity and genetic variants on PLTP activity.J Lipid Res. 2015 Jul;56(7):1351-62. doi: 10.1194/jlr.P058032. Epub 2015 May 25. J Lipid Res. 2015. PMID: 26009633 Free PMC article.
-
Dysfunctional High-Density Lipoprotein: An Innovative Target for Proteomics and Lipidomics.Cholesterol. 2015;2015:296417. doi: 10.1155/2015/296417. Epub 2015 Nov 8. Cholesterol. 2015. PMID: 26634153 Free PMC article. Review.
-
Obstructive Sleep Apnoea and Lipid Metabolism: The Summary of Evidence and Future Perspectives in the Pathophysiology of OSA-Associated Dyslipidaemia.Biomedicines. 2022 Oct 29;10(11):2754. doi: 10.3390/biomedicines10112754. Biomedicines. 2022. PMID: 36359273 Free PMC article. Review.
-
Novel Apo E-Derived ABCA1 Agonist Peptide (CS-6253) Promotes Reverse Cholesterol Transport and Induces Formation of preβ-1 HDL In Vitro.PLoS One. 2015 Jul 24;10(7):e0131997. doi: 10.1371/journal.pone.0131997. eCollection 2015. PLoS One. 2015. PMID: 26207756 Free PMC article.
-
Tweaking the cholesterol efflux capacity of reconstituted HDL.Biochem Cell Biol. 2012 Oct;90(5):636-45. doi: 10.1139/o2012-015. Epub 2012 May 18. Biochem Cell Biol. 2012. PMID: 22607224 Free PMC article.
References
-
- Brewer H. B., Santamarina-Fojo S. 2003. New insights into the role of the adenosine triphosphate-binding cassette transporters in high-density lipoprotein metabolism and reverse cholesterol transport. Am. J. Cardiol. 91: 3E–11E - PubMed
-
- Tall A. R.1993. Plasma cholesteryl ester transfer protein. J. Lipid Res. 34: 1255–1274 - PubMed
-
- Timmins J. M., Lee J. Y., Boudyguina E., Kluckman K. D., Brunham L. R., Mulya A., Gebre A. K., Cutinho J. M., Colvin P. L., Smith T. L., et al. 2005. Targeted inactivation of hepatic Abca1 causes profound hypoalphalipoproteinemia and kidney hypercatabolism of apoA-I. J. Clin. Invest. 115: 1333–1342 - PMC - PubMed
-
- McCall M. R., Nichols A. V., Blanche P. J., Shore V. G., Forte T. M. 1989. Lecithin:cholesterol acyltransferase-induce transformation of HepG2 lipoproteins. J. Lipid Res. 30: 1579–1589 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
