

Human-induced pluripotent stem cells from blood cells of healthy donors and patients with acquired blood disorders
- PMID: 19797525
- PMCID: PMC2798863
- DOI: 10.1182/blood-2009-04-217406
Human-induced pluripotent stem cells from blood cells of healthy donors and patients with acquired blood disorders
Abstract
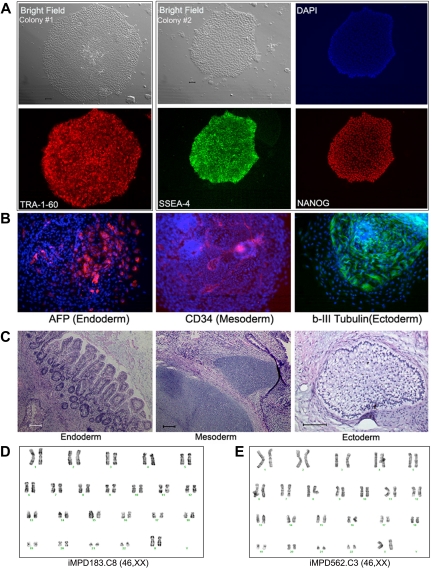
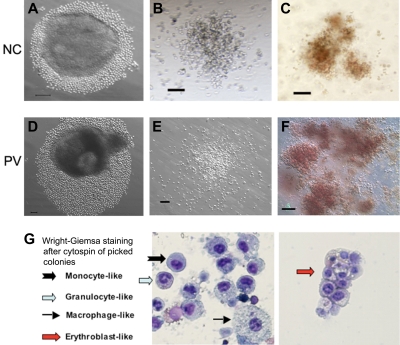
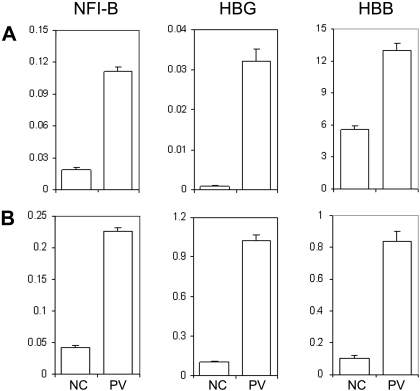
Human induced pluripotent stem (iPS) cells derived from somatic cells hold promise to develop novel patient-specific cell therapies and research models for inherited and acquired diseases. We and others previously reprogrammed human adherent cells, such as postnatal fibroblasts to iPS cells, which resemble adherent embryonic stem cells. Here we report derivation of iPS cells from postnatal human blood cells and the potential of these pluripotent cells for disease modeling. Multiple human iPS cell lines were generated from previously frozen cord blood or adult CD34(+) cells of healthy donors, and could be redirected to hematopoietic differentiation. Multiple iPS cell lines were also generated from peripheral blood CD34(+) cells of 2 patients with myeloproliferative disorders (MPDs) who acquired the JAK2-V617F somatic mutation in their blood cells. The MPD-derived iPS cells containing the mutation appeared normal in phenotypes, karyotype, and pluripotency. After directed hematopoietic differentiation, the MPD-iPS cell-derived hematopoietic progenitor (CD34(+)CD45(+)) cells showed the increased erythropoiesis and gene expression of specific genes, recapitulating features of the primary CD34(+) cells of the corresponding patient from whom the iPS cells were derived. These iPS cells provide a renewable cell source and a prospective hematopoiesis model for investigating MPD pathogenesis.
Figures


Comment in
-
Neoplastic blood cells become pluripotent.Blood. 2009 Dec 24;114(27):5409-10. doi: 10.1182/blood-2009-10-247502. Blood. 2009. PMID: 20035040 No abstract available.
References
-
- Takahashi K, Tanabe K, Ohnuki M, et al. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell. 2007;131(5):861–872. - PubMed
-
- Yu J, Vodyanik M, Smuga-Otto K, et al. Induced pluripotent stem cell lines derived from human somatic cells. Science. 2007;318(5858):1917–1920. - PubMed
-
- Park I, Zhao R, West J, et al. Reprogramming of human somatic cells to pluripotency with defined factors. Nature. 2008;451(7175):141–146. - PubMed
-
- Mali P, Ye Z, Hommond H, et al. Improved efficiency and pace of generating induced pluripotent stem cells from human adult and fetal fibroblasts. Stem Cells. 2008;26(8):1998–2005. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
