

Auxin-binding proteins without KDEL sequence in the moss Funaria hygrometrica
- PMID: 19798504
- PMCID: PMC2766497
- DOI: 10.1007/s00299-009-0775-2
Auxin-binding proteins without KDEL sequence in the moss Funaria hygrometrica
Abstract
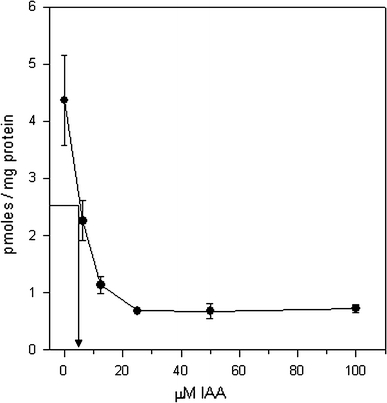
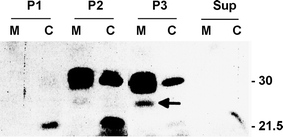
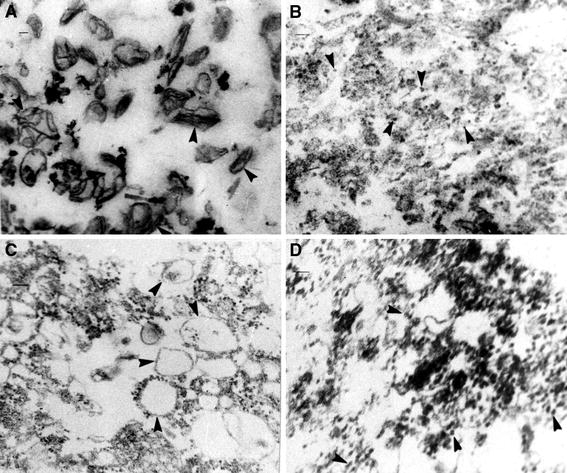
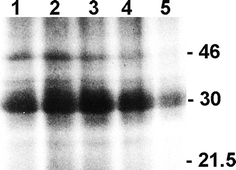
Whereas the important plant growth regulator auxin has multiple effects in flowering plants, it induces a specific cell differentiation step in the filamentous moss protonema. Here, we analyse the presence of classical auxin-binding protein (ABP1) homologues in the moss Funaria hygrometrica. Microsomal membranes isolated from protonemata of F. hygrometrica have specific indole acetic acid-binding sites, estimated to be about 3-5 pmol/mg protein with an apparent dissociation constant (K (d)) between 3 and 5 microM. Western analyses with anti-ABP1 antiserum detected the canonical endoplasmic reticulum (ER)-localised 22-24 kDa ABP1 in Zea mays, but not in F. hygrometrica. Instead, polypeptides of 31-33 and 46 kDa were labelled in the moss as well as in maize. In F. hygrometrica these proteins were found exclusively in microsomal membrane fractions and were confirmed as ABPs by photo-affinity labelling with 5-azido-[7-(3)H]-indole-3-acetic acid. Unlike the classical corn ABP1, these moss ABPs did not contain the KDEL ER retention sequence. Consistently, the fully sequenced genome of the moss Physcomitrella patens, a close relative of F. hygrometrica, encodes an ABP1-homologue without KDEL sequence. Our study suggests the presence of putative ABPs in F. hygrometrica that share immunological epitopes with ABP1 and bind auxin but are different from the classical corn ABP1.
Figures




Similar articles
-
Molecular dynamics simulations of the auxin-binding protein 1 in complex with indole-3-acetic acid and naphthalen-1-acetic acid.Proteins. 2014 Oct;82(10):2744-55. doi: 10.1002/prot.24639. Epub 2014 Jul 17. Proteins. 2014. PMID: 25043515
-
Assigning DYW-type PPR proteins to RNA editing sites in the funariid mosses Physcomitrella patens and Funaria hygrometrica.Plant J. 2011 Jul;67(2):370-80. doi: 10.1111/j.1365-313X.2011.04600.x. Epub 2011 May 11. Plant J. 2011. PMID: 21466601
-
Retention of maize auxin-binding protein in the endoplasmic reticulum: quantifying escape and the role of auxin.Planta. 1997;202(3):313-23. doi: 10.1007/s004250050133. Planta. 1997. PMID: 9232903
-
Caulonema differentiation in Funaria protonema.Int J Dev Biol. 2020;64(1-2-3):21-28. doi: 10.1387/ijdb.190290mj. Int J Dev Biol. 2020. PMID: 32659008 Review.
-
Auxin binding protein: curiouser and curiouser.Trends Plant Sci. 2001 Dec;6(12):586-90. doi: 10.1016/s1360-1385(01)02150-1. Trends Plant Sci. 2001. PMID: 11738384 Review.
Cited by
-
AUXIN BINDING PROTEIN1: the outsider.Plant Cell. 2011 Jun;23(6):2033-43. doi: 10.1105/tpc.111.087064. Epub 2011 Jun 30. Plant Cell. 2011. PMID: 21719690 Free PMC article. Review.
-
Microorganism and filamentous fungi drive evolution of plant synapses.Front Cell Infect Microbiol. 2013 Aug 15;3:44. doi: 10.3389/fcimb.2013.00044. eCollection 2013. Front Cell Infect Microbiol. 2013. PMID: 23967407 Free PMC article. Review.
References
-
- Andres DA, Dickerson IM, Dixon JE. Variants of the carboxyl-terminal KDEL sequence direct intracellular retention. J Biol Chem. 1990;265:5952–5955. - PubMed
-
- Barbier-Brygoo H, Ephritikhine G, Maurel C, Guern J. Perception of the auxin signal at the plasma membrane of tobacco mesophyll protoplasts. Biochem Soc Trans. 1992;20:59–63. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
