

Role of intracellular polysaccharide in persistence of Streptococcus mutans
- PMID: 19801415
- PMCID: PMC2786568
- DOI: 10.1128/JB.00425-09
Role of intracellular polysaccharide in persistence of Streptococcus mutans
Abstract
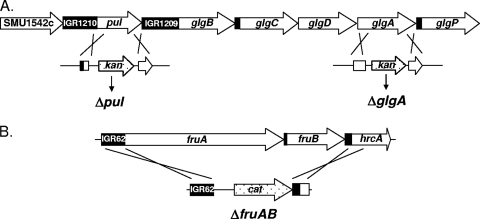
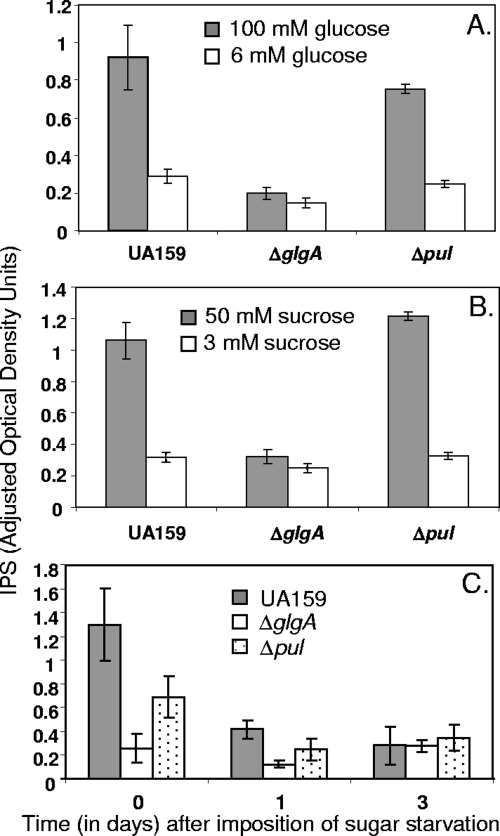
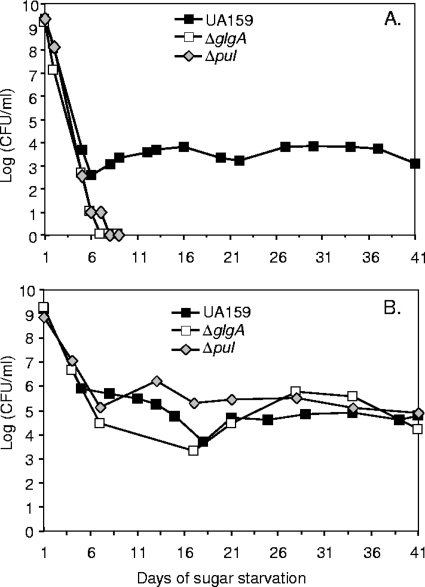
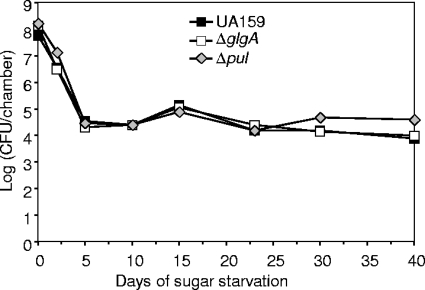
Intracellular polysaccharide (IPS) is accumulated by Streptococcus mutans when the bacteria are grown in excess sugar and can contribute toward the cariogenicity of S. mutans. Here we show that inactivation of the glgA gene (SMU1536), encoding a putative glycogen synthase, prevented accumulation of IPS. IPS is important for the persistence of S. mutans grown in batch culture with excess glucose and then starved of glucose. The IPS was largely used up within 1 day of glucose starvation, and yet survival of the parental strain was extended by at least 15 days beyond that of a glgA mutant; potentially, some feature of IPS metabolism distinct from providing nutrients is important for persistence. IPS was not needed for persistence when sucrose was the carbon source or when mucin was present.
Figures
References
-
- Carlsson, J. 1997. Bacterial metabolism in dental biofilms. Adv. Dent. Res. 11:75-80. - PubMed
-
- Chary, V. K., M. Busuioc, J. A. Renye, Jr., and P. J. Piggot. 2005. Vectors that facilitate the replacement of transcriptional lacZ fusions in Streptococcus mutans and Bacillus subtilis with fusions to gfp or gusA. FEMS Microbiol. Lett. 247:171-176. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
