

An endogenous positively selecting peptide enhances mature T cell responses and becomes an autoantigen in the absence of microRNA miR-181a
- PMID: 19801983
- PMCID: PMC3762483
- DOI: 10.1038/ni.1797
An endogenous positively selecting peptide enhances mature T cell responses and becomes an autoantigen in the absence of microRNA miR-181a
Abstract
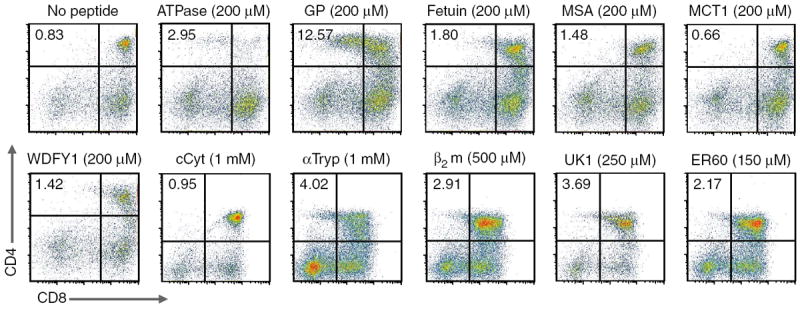
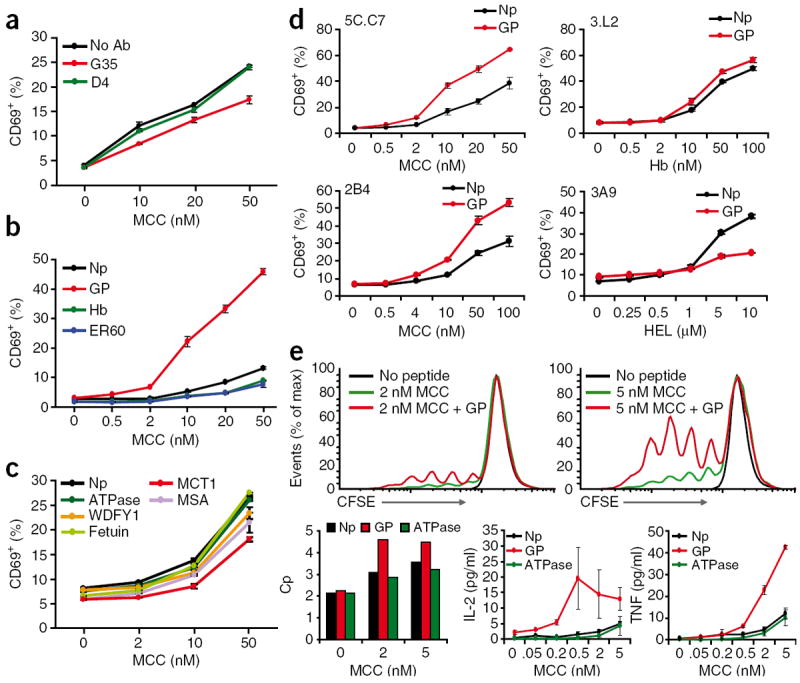
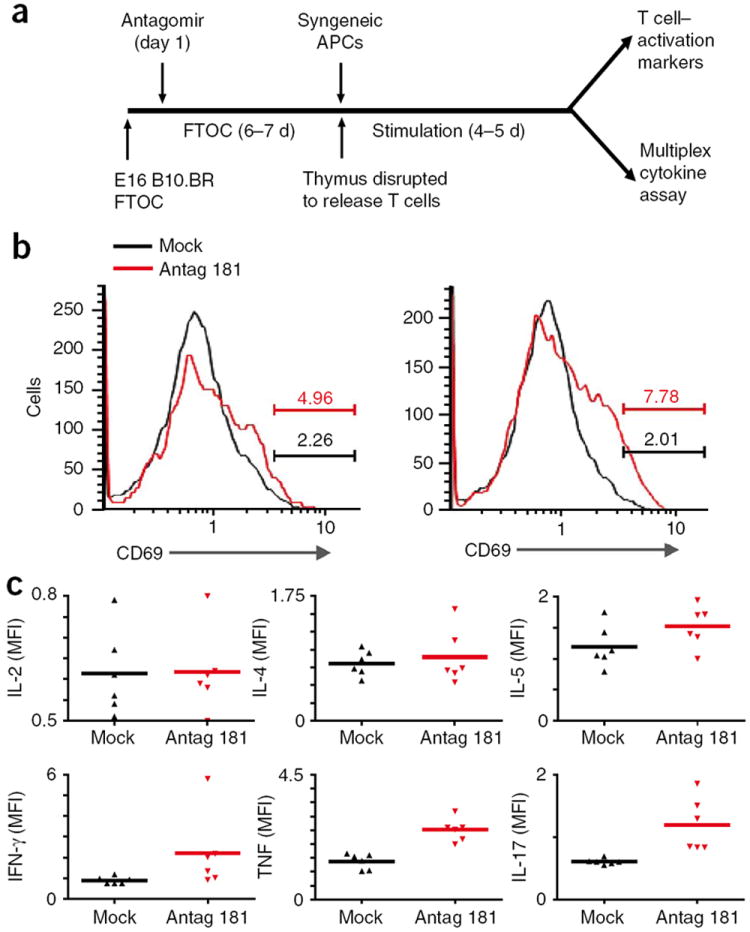
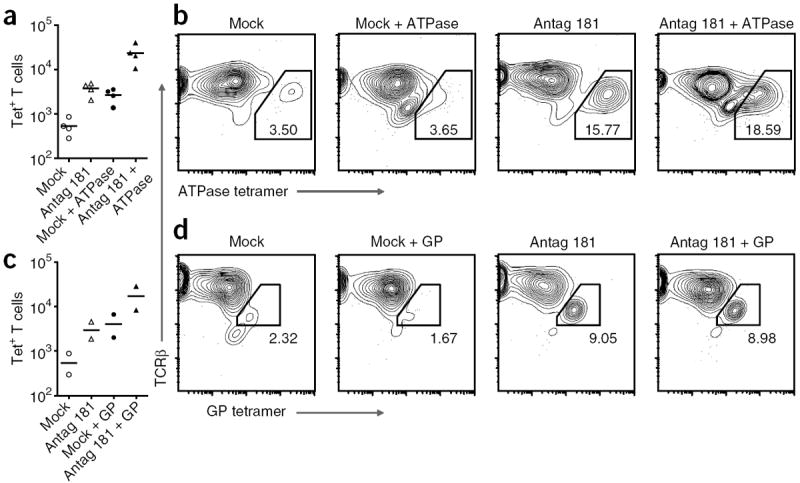
Thymic positive selection is based on the interactions of T cell antigen receptors (TCRs) with self peptide-major histocompatibility complex (MHC) ligands, but the identity of selecting peptides for MHC class II-restricted TCRs and the functional consequences of this peptide specificity are not clear. Here we identify several endogenous self peptides that positively selected the MHC class II-restricted 5C.C7 TCR. The most potent of these also enhanced mature T cell activation, which supports the hypothesis that one function of positive selection is to produce T cells that can use particular self peptide-MHC complexes for activation and/or homeostasis. We also show that inhibiting the microRNA miR-181a resulted in maturation of T cells that overtly reacted toward these erstwhile positively selecting peptides. Therefore, miR-181a helps to guarantee the clonal deletion of particular moderate-affinity clones by modulating the TCR signaling threshold of thymocytes.
Conflict of interest statement
COMPETING INTERESTS STATEMENT
The authors declare competing financial interests: details accompany the full-text HTML version of the paper at
Figures
Comment in
-
Positively selecting peptides: their job does not end in the thymus.Nat Immunol. 2009 Nov;10(11):1143-4. doi: 10.1038/ni1109-1143. Nat Immunol. 2009. PMID: 19841646
References
-
- Kisielow P, Bluthmann H, Staerz UD, Steinmetz M, von Boehmer H. Tolerance in T-cell-receptor transgenic mice involves deletion of nonmature CD4+8+ thymocytes. Nature. 1988;333:742–746. - PubMed
-
- Ernst B, Lee DS, Chang JM, Sprent J, Surh CD. The peptide ligands mediating positive selection in the thymus control T cell survival and homeostatic proliferation in the periphery. Immunity. 1999;11:173–181. - PubMed
-
- Stefanova I, Dorfman JR, Tsukamoto M, Germain RN. On the role of self-recognition in T cell responses to foreign antigen. Immunol Rev. 2003;191:97–106. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
