

Protein-mediated molecular bridging: a key mechanism in biopolymer organization
- PMID: 19804731
- PMCID: PMC2756379
- DOI: 10.1016/j.bpj.2009.06.051
Protein-mediated molecular bridging: a key mechanism in biopolymer organization
Abstract
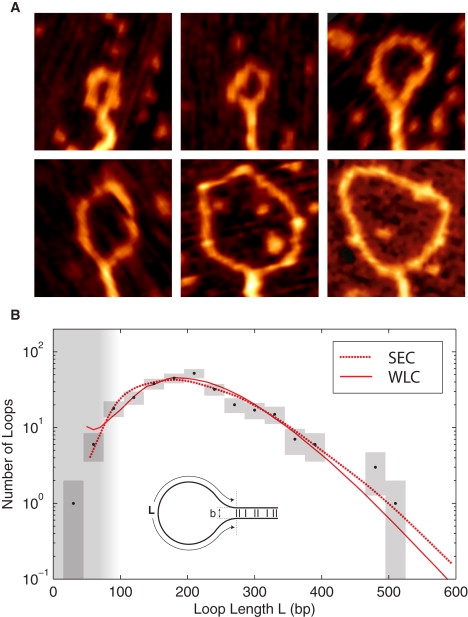
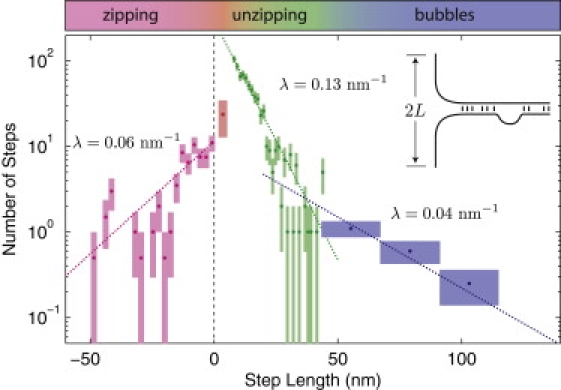
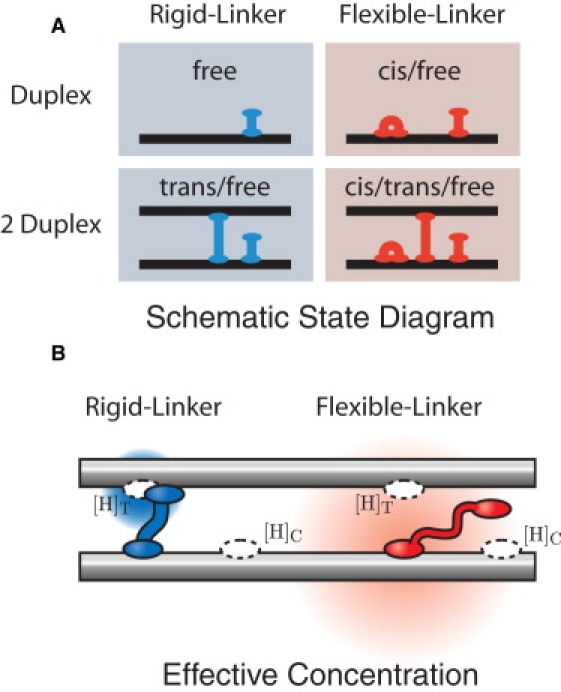
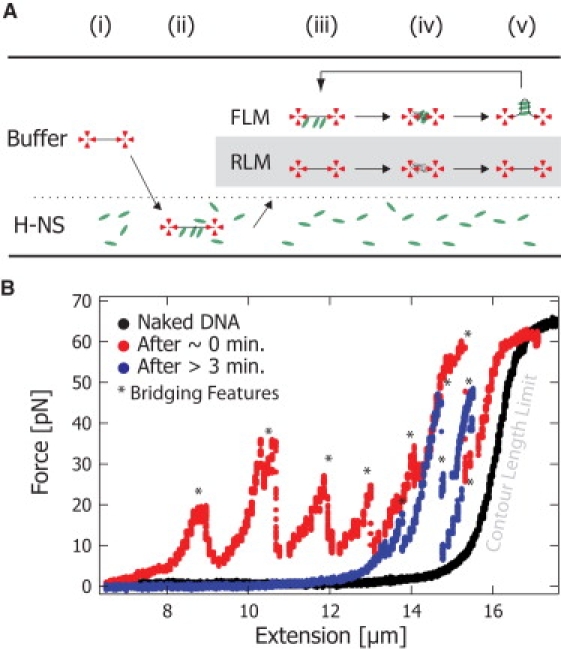
Protein-mediated bridging is ubiquitous and essential for shaping cellular structures in all organisms. Here we dissect this mechanism for a model system: the Histone-like Nucleoid-Structuring protein (H-NS). We present data from two complementary single-molecule assays that probe the H-NS-DNA interaction: a dynamic optical-trap-driven unzipping assay and an equilibrium H-NS-mediated DNA looping scanning force microscopy imaging assay. To quantitatively analyze and compare these assays, we employ what we consider a novel theoretical framework that describes the bridging motif. The interplay between the experiments and our theoretical model not only infers the effective interaction free energy, the bridging conformation and the duplex-duplex spacing, but also reveals a second, unresolved, cis-binding mode that challenges our current understanding of the role of bridging proteins in chromatin structure. We expect that this theoretical framework for describing protein-mediated bridging will be applicable to proteins acting in chromatin and cytoskeletal organization.
Figures
References
-
- Dorman C.J. H-NS: a universal regulator for a dynamic genome. Nat. Rev. Microbiol. 2004;2:391–400. - PubMed
-
- Dorman C.J. H-NS, the genome sentinel. Nat. Rev. Microbiol. 2007;5:157–161. - PubMed
-
- Dame R.T., Noom M.C., Wuite G.J.L. Bacterial chromatin organization by H-NS protein unravelled using dual DNA manipulation. Nature. 2006;444:387–390. - PubMed
-
- Dame R.T. The role of nucleoid-associated proteins in the organization and compaction of bacterial chromatin. Mol. Microbiol. 2005;56:858–870. - PubMed
-
- Luijsterburg M.S., Noom M.C., Wuite G.J.L., Dame R.T. The architectural role of nucleoid-associated proteins in the organization of bacterial chromatin: a molecular perspective. J. Struct. Biol. 2006;156:262–272. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
