

Review
doi: 10.1016/j.cell.2009.09.015.
Olfactory perception: receptors, cells, and circuits
Affiliations
- PMID: 19804753
- PMCID: PMC2765334
- DOI: 10.1016/j.cell.2009.09.015
Item in Clipboard
Review
Olfactory perception: receptors, cells, and circuits
Cell.
.
Abstract
Remarkable advances in our understanding of olfactory perception have been made in recent years, including the discovery of new mechanisms of olfactory signaling and new principles of olfactory processing. Here, we discuss the insight that has been gained into the receptors, cells, and circuits that underlie the sense of smell.
Figures

(A) Sagittal view of a rodent head, showing four olfactory organs: the main olfactory epithelium (MOE), vomeronasal organ (VNO), Grueneberg ganglion (GG) and septal organ of Masera (SO). Olfactory receptor neurons (ORNs) in the MOE, GG and SO all project to the main olfactory bulb (MOB), whereas the VNO neurons project to the accessory olfactory bulb (AOB). Olfactory information is further processed in higher brain regions, such as the anterior olfactory nucleus (AON), the olfactory tubercle (OT), entorhinal cortex (ENT), piriform cortex (PIR) and cortical amygdala (CoAMG). Inset: coronal section of the brain. (B) Frontal view of a Drosophila head. There are two pairs of olfactory organs: the third antennal segments and maxillary palps. Olfactory information is first relayed to the antennal lobe, which contains multiple glomeruli. Subsequent processing takes place at the lateral horn of the protocerebrum and Kenyon cells in the mushroom body. Connectivity has been simplified for clarity.

Excitatory neurons are shown in orange and inhibitory neurons in blue. (A) Olfactory receptor neurons (ORNs) in the olfactory epithelium that express different olfactory receptors project axons to separate glomeruli (dashed outlines) in the olfactory bulb where they synapse on mitral and tufted (M/T) cells, whose apical dendrite is usually localized to a single glomerulus. Juxtaglomerular cells (blue) contribute to intraglomerular inhibition. In the glomerulus, ORNs form synapses on juxtaglomerular cell dendrites, which in turn inhibit ORN axon terminals. Reciprocal synapses are also found between juxtaglomerular cell and M/T cell dendrites. Reciprocal synapses are formed between the dendrites of granule cells and M/T cells. M/T cells excite granule cells, which respond by inhibiting M/T cells. Due to the lateral spread of M/T secondary dendrites, granule cells contact multiple M/T cells associated with different glomeruli, and thus can mediate both intra- and interglomerular inhibition. (B) In Drosophila, ORNs expressing the same olfactory receptors in the antenna or maxillary palp synapse on projection neurons in a single glomerulus, analogous to the olfactory bulb. GABA-releasing local neurons (LNs) pre-synaptically inhibit ORN axon terminals in multiple glomeruli, mediating interglomerular inhibition. Excitatory cholinergic LNs mediate interglomerular excitation.

(A) Responses of 48 second-order neurons (projection neurons, PNs) and 42 third-order neurons (Kenyon cells) in locusts to a panel of 14 odors. Black squares indicate activation of a neuron by an odor; white squares indicate lack of response or inhibitory responses. Ns respond to many odors, in contrast to Kenyon cells, which respond to only a few (sparse coding). Additionally, similar activation patterns of 2nd order neurons, e.g. odors 7 (blue) and 13 (red), result in highly divergent activation patterns of 3rd order neurons, a process termed “decorrelation”. This decorrelation is thought to make these odors easier to discriminate. Adapted from Perez-Orive et al., 2002. Reprinted with permission from AAAS. (B) A simplified illustration of how sparsening and decorrelation of responses occur in 3rd order neurons. Odors A and B each activate multiple 2nd order neurons (colored circles), with similar patterns of activated neurons. However, due to the requirement of synchronized inputs from multiple 2nd order neurons (coincidence detection), many fewer 3rd order neurons are activated (sparsening) with more distinct activation pattern (decorrelation).
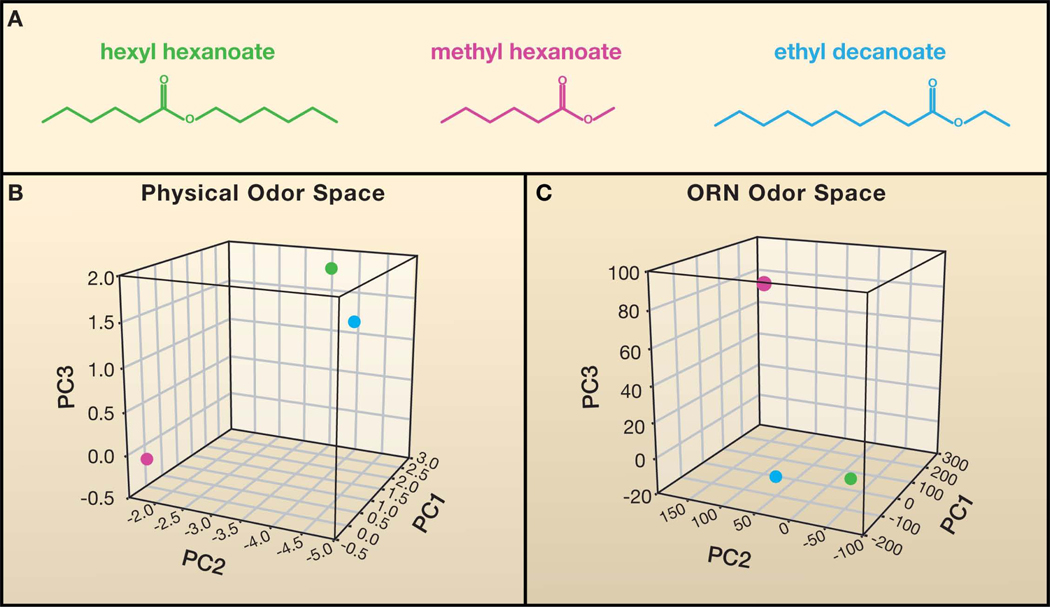
(A) Chemical structures of three odorants. It is difficult to compare the degree of relatedness between odorants by visual inspection. (B) A physical odor space constructed using 32 optimized descriptors of odorant structure (Haddad et al., 2008), for the odor panel used in Hallem and Carlson, 2006. The Euclidean distance between hexyl hexanoate (green) and ethyl decanoate (blue) is smaller than the distance of either odorant to methyl hexanoate (magenta), indicating that hexyl hexanoate and ethyl decanoate are more structurally similar. The first three principal components (PC) are shown. Similarly, hexyl hexanoate and ethyl decanoate map closer to each other than to methyl hexanoate in a neural odor space (C), based on the functional data reported in Hallem and Carlson, 2006.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
