

Structure of an integrin alphaIIb beta3 transmembrane-cytoplasmic heterocomplex provides insight into integrin activation
- PMID: 19805198
- PMCID: PMC2764936
- DOI: 10.1073/pnas.0909589106
Structure of an integrin alphaIIb beta3 transmembrane-cytoplasmic heterocomplex provides insight into integrin activation
Abstract
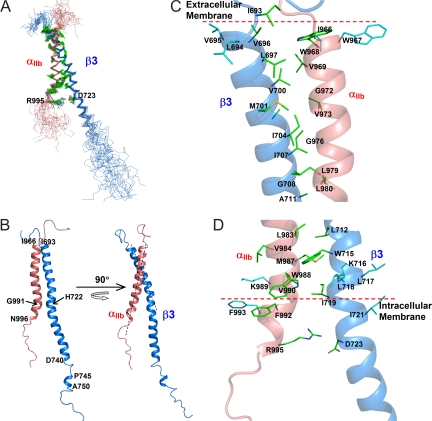
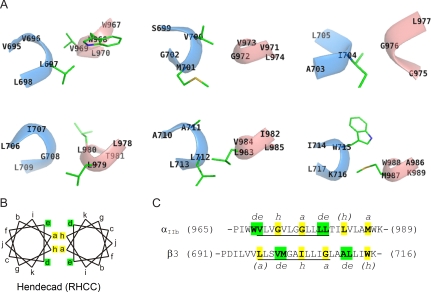
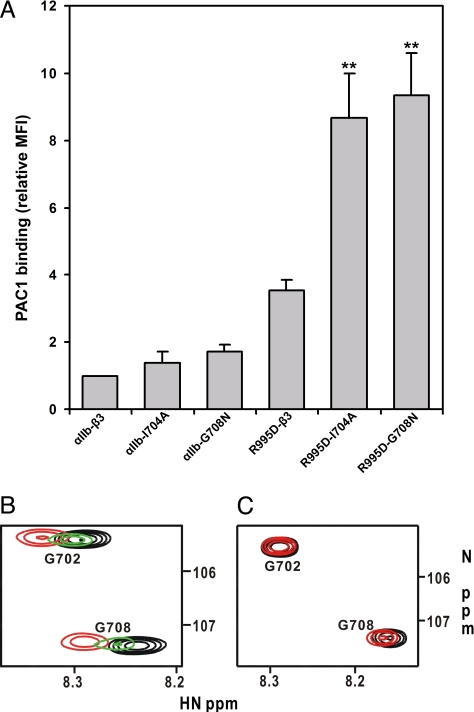
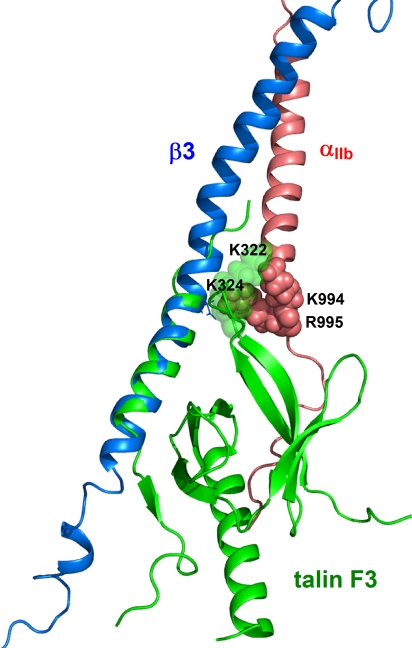
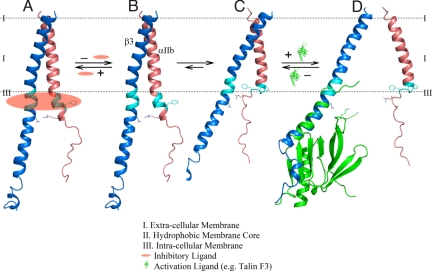
Heterodimeric integrin adhesion receptors regulate diverse biological processes including angiogenesis, thrombosis and wound healing. The transmembrane-cytoplasmic domains (TMCDs) of integrins play a critical role in controlling activation of these receptors via an inside-out signaling mechanism, but the precise structural basis remains elusive. Here, we present the solution structure of integrin alphaIIb beta3 TMCD heterodimer, which reveals a right-handed coiled-coil conformation with 2 helices intertwined throughout the transmembrane region. The helices extend into the cytoplasm and form a clasp that differs significantly from a recently published alphaIIb beta3 TMCD structure. We show that while a point mutation in the clasp interface modestly activates alphaIIb beta3, additional mutations in the transmembrane interface have a synergistic effect, leading to extensive integrin activation. Detailed analyses and structural comparison with previous studies suggest that extensive integrin activation is a highly concerted conformational transition process, which involves transmembrane coiled-coil unwinding that is triggered by the membrane-mediated alteration and disengagement of the membrane-proximal clasp. Our results provide atomic insight into a type I transmembrane receptor heterocomplex and the mechanism of integrin inside-out transmembrane signaling.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
