

Limited terminal transferase in human DNA polymerase mu defines the required balance between accuracy and efficiency in NHEJ
- PMID: 19805281
- PMCID: PMC2738622
- DOI: 10.1073/pnas.0908492106
Limited terminal transferase in human DNA polymerase mu defines the required balance between accuracy and efficiency in NHEJ
Abstract
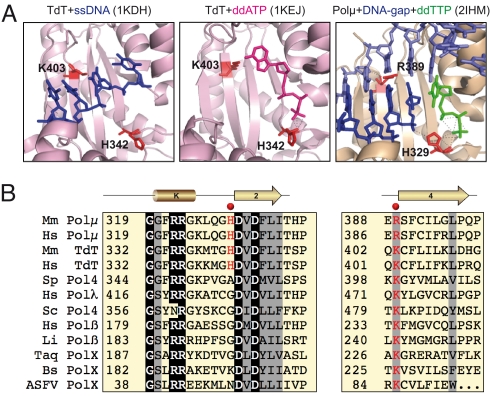
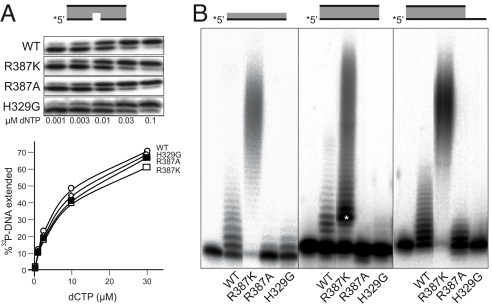
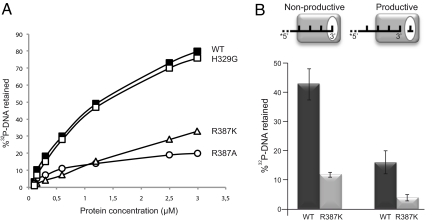
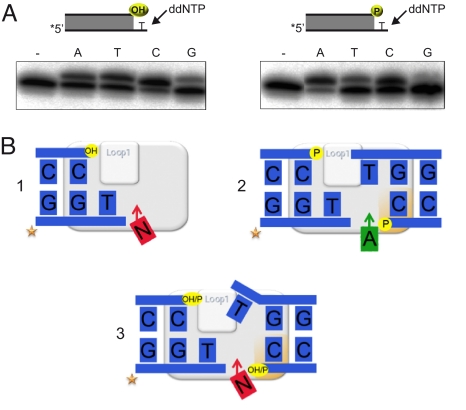
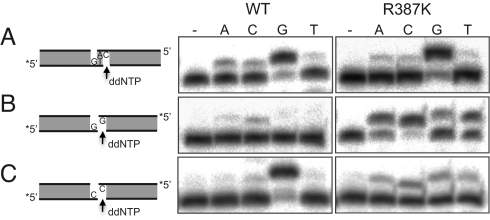
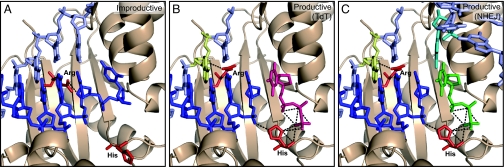
DNA polymerase mu (Polmu) is a family X member implicated in DNA repair, with template-directed and terminal transferase (template-independent) activities. It has been proposed that the terminal transferase activity of Polmu can be specifically required during non-homologous end joining (NHEJ) to create or increase complementarity of DNA ends. By site-directed mutagenesis in human Polmu, we have identified a specific DNA ligand residue (Arg(387)) that is responsible for its limited terminal transferase activity compared to that of human TdT, its closest homologue (42% amino acid identity). Polmu mutant R387K (mimicking TdT) displayed a large increase in terminal transferase activity, but a weakened interaction with ssDNA. That paradox can be explained by the regulatory role of Arg(387) in the translocation of the primer from a non-productive E:DNA complex to a productive E:DNA:dNTP complex in the absence of a templating base, which is probably the rate limiting step during template-independent synthesis. Further, we show that the Polmu switch from terminal transferase to templated insertions in NHEJ reactions is triggered by recognition of a 5'-P at a second DNA end, whose 3'-protrusion could provide a templating base to facilitate such a special "pre-catalytic translocation step." These studies shed light on the mechanism by which a rate-limited terminal transferase activity in Polmu could regulate the balance between accuracy and necessary efficiency, providing some variability during NHEJ.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Friedberg EC, et al. DNA Repair and Mutagenesis. 2nd Ed. Washington D.C.: ASM Press; 2006.
-
- Bebenek K, Kunkel TA. Functions of DNA polymerases. Adv Protein Chem. 2004;69:137–165. - PubMed
-
- Bork P, et al. A superfamily of conserved domains in DNA damage-responsive cell cycle checkpoint proteins. FASEB J. 1997;11:68–76. - PubMed
-
- Callebaut I, Mornon JP. From BRCA1 to RAP1: A widespread BRCT module closely associated with DNA repair. FEBS Lett. 1997;400:25–30. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
