

Experience-dependent maturation of the glomerular microcircuit
- PMID: 19805387
- PMCID: PMC2757847
- DOI: 10.1073/pnas.0808946106
Experience-dependent maturation of the glomerular microcircuit
Abstract
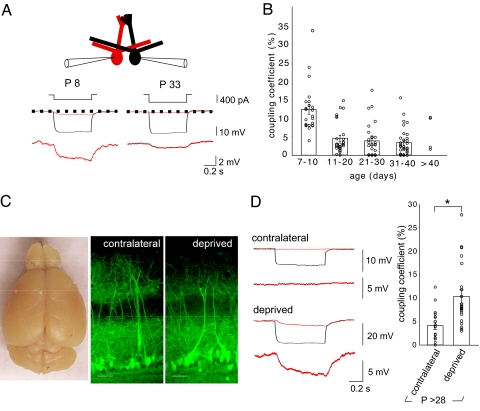
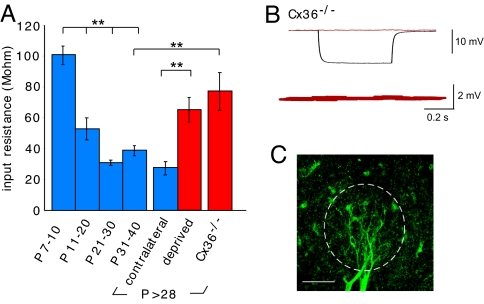
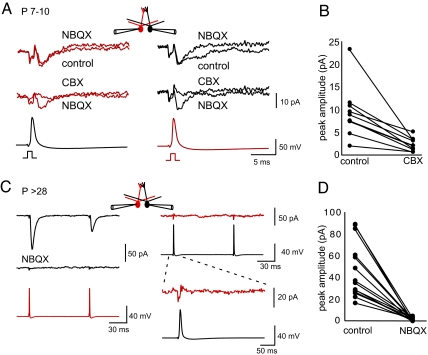
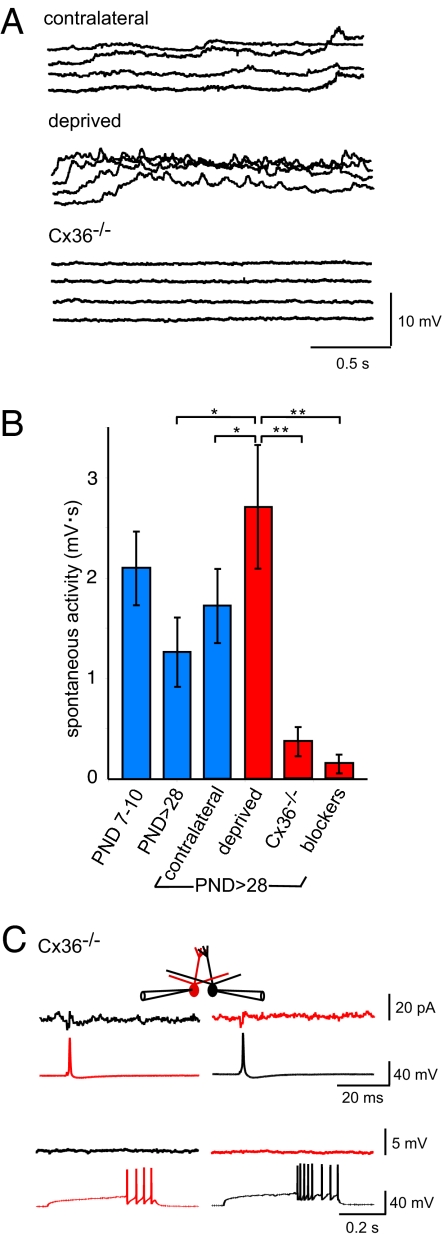
Spontaneous and patterned activity, largely attributed to chemical transmission, shape the development of virtually all neural circuits. However, electrical transmission also has an important role in coordinated activity in the brain. In the olfactory bulb, gap junctions between apical dendrites of mitral cells increase excitability and synchronize firing within each glomerulus. We report here that the development of the glomerular microcircuit requires both sensory experience and connexin (Cx)36-mediated gap junctions. Coupling coefficients, which measure electrical coupling between mitral cell dendrites, were high in young mice, but decreased after postnatal day (P)10 because of a maturational increase in membrane conductance. Sensory deprivation, induced by unilateral naris occlusion at birth, slowed the morphological development of mitral cells and arrested the maturational changes in membrane conductance and coupling coefficients. As the coupling coefficients decreased in normal mice, a glutamate-mediated excitatory postsynaptic current (EPSC) between mitral cells emerged by P30. Although mitral-mitral EPSCs were generally unidirectional, they were not present in young adult Cx36(-/-) mice, suggesting that gap junctions are required for the development and/or function of the mature circuit. The experience-dependent transition from electrical transmission to combined chemical and electrical transmission provides a previously unappreciated mechanism that may tune the response properties of the glomerular microcircuit.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Katz LC, Shatz CJ. Synaptic activity and the construction of cortical circuits. Science. 1996;274:1133–1138. - PubMed
-
- Grubb MS, Thompson ID. The influence of early experience on the development of sensory systems. Curr Opin Neurobiol. 2004;14:503–512.51. - PubMed
-
- Fox K, Wong RO. A comparison of experience-dependent plasticity in the visual and somatosensory systems. Neuron. 2005;48:465–477. - PubMed
-
- Bennett MVL, et al. Gap junctions: New tools, new answers, new questions. Neuron. 1991;6:305–320. - PubMed
-
- Roerig B, Feller MB. Neurotransmitters and gap junctions in developing neural circuits. Brain Res Brain Res Rev. 2000;32:86–114. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
