

Transforming growth factor beta is dispensable for the molecular orchestration of Th17 cell differentiation
- PMID: 19808254
- PMCID: PMC2768861
- DOI: 10.1084/jem.20082286
Transforming growth factor beta is dispensable for the molecular orchestration of Th17 cell differentiation
Erratum in
- J Exp Med. 2009 Dec 21;206(13):3157
Abstract
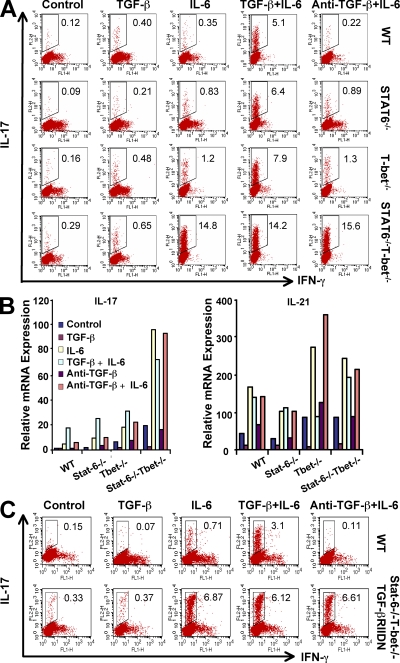
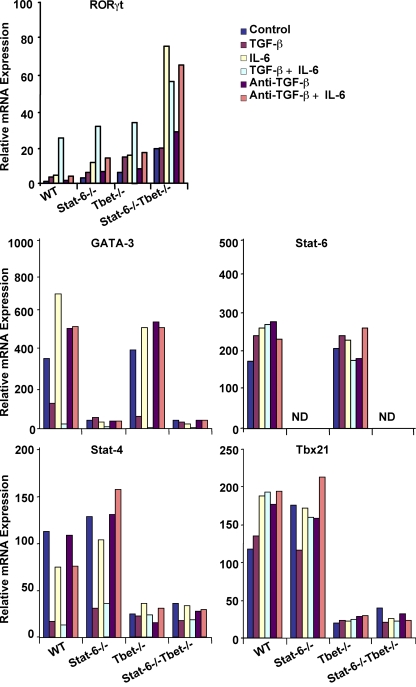
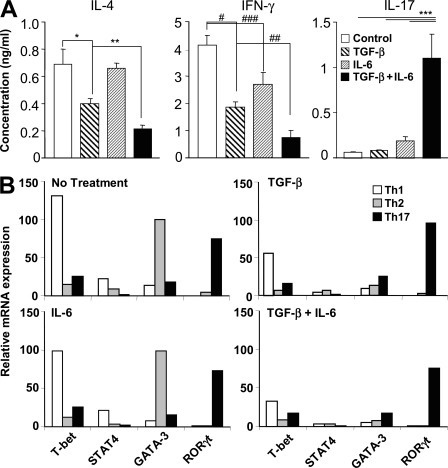
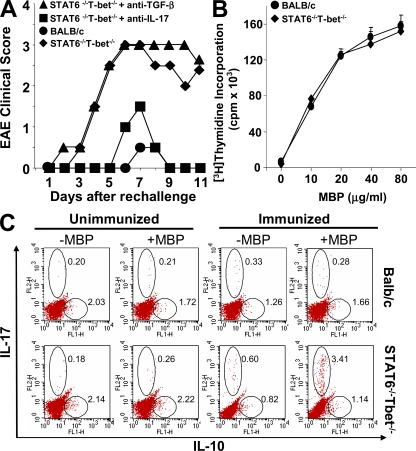
Interleukin (IL)-17-producing T helper (Th17) cells play a critical role in the pathophysiology of several autoimmune disorders. The differentiation of Th17 cells requires the simultaneous presence of an unusual combination of cytokines: IL-6, a proinflammatory cytokine, and transforming growth factor (TGF) beta, an antiinflammatory cytokine. However, the molecular mechanisms by which TGF-beta exerts its effects on Th17 cell differentiation remain elusive. We report that TGF-beta does not directly promote Th17 cell differentiation but instead acts indirectly by blocking expression of the transcription factors signal transducer and activator of transcription (STAT) 4 and GATA-3, thus preventing Th1 and Th2 cell differentiation. In contrast, TGF-beta had no effect on the expression of retinoic acid receptor-related orphan nuclear receptor gammat, a Th17-specific transcription factor. Interestingly, in Stat-6(-/-)T-bet(-/-) mice, which are unable to generate Th1 and Th2 cells, IL-6 alone was sufficient to induce robust differentiation of Th17 cells, whereas TGF-beta had no effect, suggesting that TGF-beta is dispensable for Th17 cell development. Consequently, BALB/c Stat-6(-/-)T-bet(-/-) mice, but not wild-type BALB/c mice, were highly susceptible to the development of experimental autoimmune encephalomyelitis, which could be blocked by anti-IL-17 antibodies but not by anti-TGF-beta antibodies. Collectively, these data provide evidence that TGF-beta is not directly required for the molecular orchestration of Th17 cell differentiation.
Figures
References
-
- Burlingham W.J., Love R.B., Jankowska-Gan E., Haynes L.D., Xu Q., Bobadilla J.L., Meyer K.C., Hayney M.S., Braun R.K., Greenspan D.S., et al. 2007. IL-17-dependent cellular immunity to collagen type V predisposes to obliterative bronchiolitis in human lung transplants. J. Clin. Invest. 117:3498–3506 10.1172/JCI28031 - DOI - PMC - PubMed
-
- Choy E.H., Isenberg D.A., Garrood T., Farrow S., Ioannou Y., Bird H., Cheung N., Williams B., Hazleman B., Price R., et al. 2002. Therapeutic benefit of blocking interleukin-6 activity with an anti-interleukin-6 receptor monoclonal antibody in rheumatoid arthritis: a randomized, double-blind, placebo-controlled, dose-escalation trial. Arthritis Rheum. 46:3143–3150 10.1002/art.10623 - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
