

Production of beta-globin and adult hemoglobin following G418 treatment of erythroid precursor cells from homozygous beta(0)39 thalassemia patients
- PMID: 19810011
- PMCID: PMC3572903
- DOI: 10.1002/ajh.21539
Production of beta-globin and adult hemoglobin following G418 treatment of erythroid precursor cells from homozygous beta(0)39 thalassemia patients
Abstract
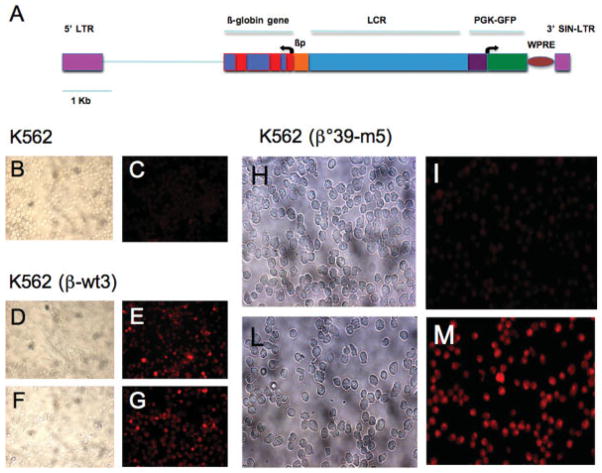
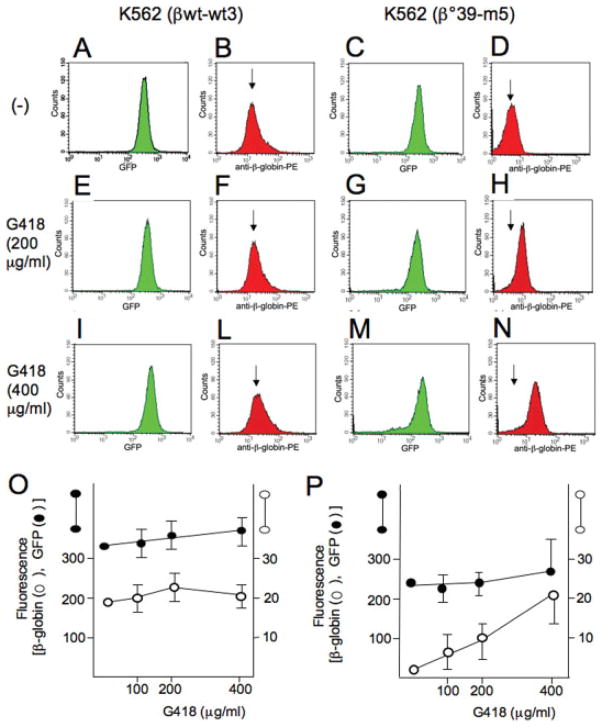
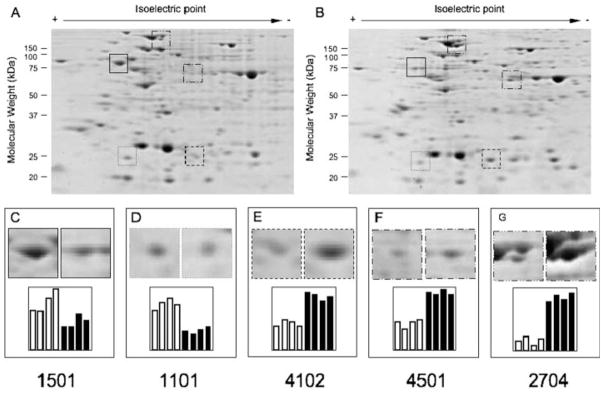
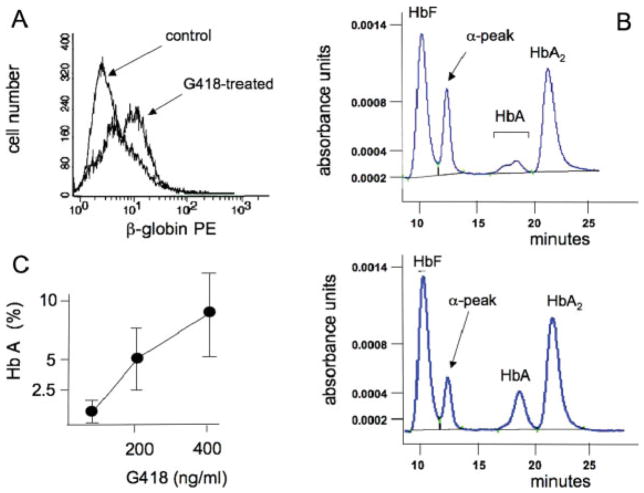
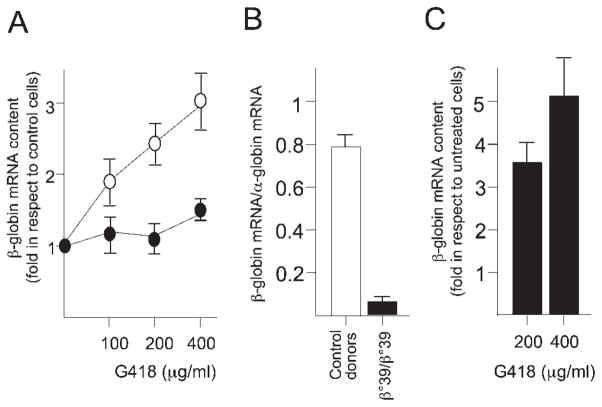
In several types of thalassemia (including beta(0)39-thalassemia), stop codon mutations lead to premature translation termination and to mRNA destabilization through nonsense-mediated decay. Drugs (for instance aminoglycosides) can be designed to suppress premature termination, inducing a ribosomal readthrough. These findings have introduced new hopes for the development of a pharmacologic approach to the cure of this disease. However, the effects of aminoglycosides on globin mRNA carrying beta-thalassemia stop mutations have not yet been investigated. In this study, we have used a lentiviral construct containing the beta(0)39-thalassemia globin gene under control of the beta-globin promoter and a LCR cassette. We demonstrated by fluorescence-activated cell sorting (FACS) analysis the production of beta-globin by K562 cell clones expressing the beta(0)39-thalassemia globin gene and treated with G418. More importantly, after FACS and high-performance liquid chromatography (HPLC) analyses, erythroid precursor cells from beta(0)39-thalassemia patients were demonstrated to be able to produce beta-globin and adult hemoglobin after treatment with G418. This study strongly suggests that ribosomal readthrough should be considered a strategy for developing experimental strategies for the treatment of beta(0)-thalassemia caused by stop codon mutations. Am. J. Hematol., 2009. (c) 2009 Wiley-Liss, Inc.
Conflict of interest statement
Conflict of interest: Nothing to report.
Figures
Similar articles
-
Screening Readthrough Compounds to Suppress Nonsense Mutations: Possible Application to β-Thalassemia.J Clin Med. 2020 Jan 21;9(2):289. doi: 10.3390/jcm9020289. J Clin Med. 2020. PMID: 31972957 Free PMC article. Review.
-
Development of K562 cell clones expressing beta-globin mRNA carrying the beta039 thalassaemia mutation for the screening of correctors of stop-codon mutations.Biotechnol Appl Biochem. 2009 Jul 9;54(1):41-52. doi: 10.1042/BA20080266. Biotechnol Appl Biochem. 2009. PMID: 19216718 Free PMC article.
-
A combined approach for β-thalassemia based on gene therapy-mediated adult hemoglobin (HbA) production and fetal hemoglobin (HbF) induction.Ann Hematol. 2012 Aug;91(8):1201-13. doi: 10.1007/s00277-012-1430-5. Epub 2012 Mar 31. Ann Hematol. 2012. PMID: 22460946 Free PMC article.
-
Therapeutic hemoglobin levels after gene transfer in β-thalassemia mice and in hematopoietic cells of β-thalassemia and sickle cells disease patients.PLoS One. 2012;7(3):e32345. doi: 10.1371/journal.pone.0032345. Epub 2012 Mar 27. PLoS One. 2012. PMID: 22479321 Free PMC article.
-
Molecular Basis and Genetic Modifiers of Thalassemia.Hematol Oncol Clin North Am. 2018 Apr;32(2):177-191. doi: 10.1016/j.hoc.2017.11.003. Hematol Oncol Clin North Am. 2018. PMID: 29458725 Review.
Cited by
-
Screening Readthrough Compounds to Suppress Nonsense Mutations: Possible Application to β-Thalassemia.J Clin Med. 2020 Jan 21;9(2):289. doi: 10.3390/jcm9020289. J Clin Med. 2020. PMID: 31972957 Free PMC article. Review.
-
New in Vitro Assay Measuring Direct Interaction of Nonsense Suppressors with the Eukaryotic Protein Synthesis Machinery.ACS Med Chem Lett. 2018 Nov 21;9(12):1285-1291. doi: 10.1021/acsmedchemlett.8b00472. eCollection 2018 Dec 13. ACS Med Chem Lett. 2018. PMID: 30613341 Free PMC article.
-
Synthetic aminoglycosides efficiently suppress cystic fibrosis transmembrane conductance regulator nonsense mutations and are enhanced by ivacaftor.Am J Respir Cell Mol Biol. 2014 Apr;50(4):805-16. doi: 10.1165/rcmb.2013-0282OC. Am J Respir Cell Mol Biol. 2014. PMID: 24251786 Free PMC article.
-
Rescue of non-sense mutated p53 tumor suppressor gene by aminoglycosides.Nucleic Acids Res. 2011 Apr;39(8):3350-62. doi: 10.1093/nar/gkq1277. Epub 2010 Dec 10. Nucleic Acids Res. 2011. PMID: 21149266 Free PMC article.
-
Recent advances in gene therapy for thalassemia.J Pharm Bioallied Sci. 2012 Jul;4(3):194-201. doi: 10.4103/0975-7406.99020. J Pharm Bioallied Sci. 2012. PMID: 22923960 Free PMC article.
References
-
- Kondrashov AS. Direct estimates of human per nucleotide mutation rates at 20 loci causing Mendelian diseases. Hum Mutat. 2003;21:12–27. - PubMed
-
- Inácio A, Silva AL, Pinto J, et al. Nonsense mutations in close proximity to the initiation codon fail to trigger full nonsense-mediated mRNA decay. J Biol Chem. 2004;279:32170–32180. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources