

The end of a myth: cloning and characterization of the ovine melatonin MT(2) receptor
- PMID: 19814723
- PMCID: PMC2782334
- DOI: 10.1111/j.1476-5381.2009.00453.x
The end of a myth: cloning and characterization of the ovine melatonin MT(2) receptor
Abstract
Background and purpose: For many years, it was suspected that sheep expressed only one melatonin receptor (closely resembling MT(1) from other mammal species). Here we report the cloning of another melatonin receptor, MT(2), from sheep.
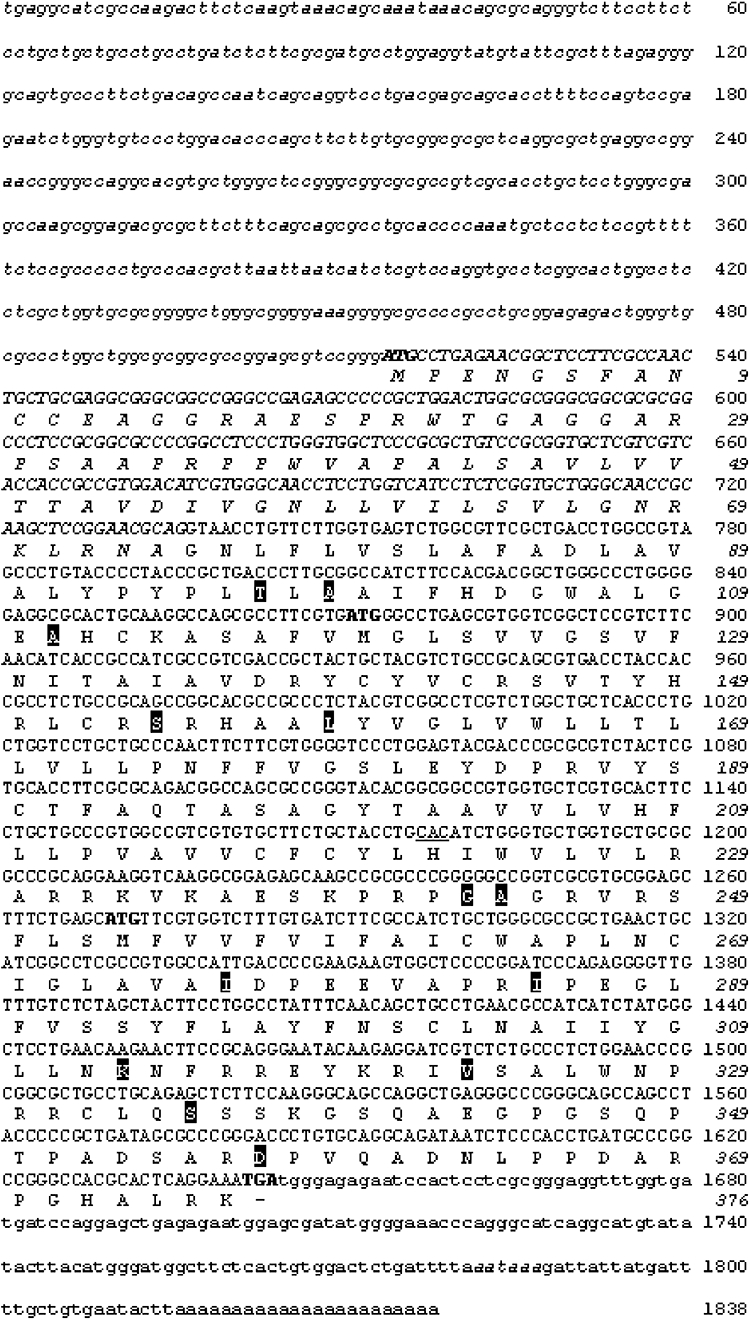
Experimental approach: Using a thermo-resistant reverse transcriptase and polymerase chain reaction primer set homologous to the bovine MT(2) mRNA sequence, we have cloned and characterized MT(2) receptors from sheep retina.
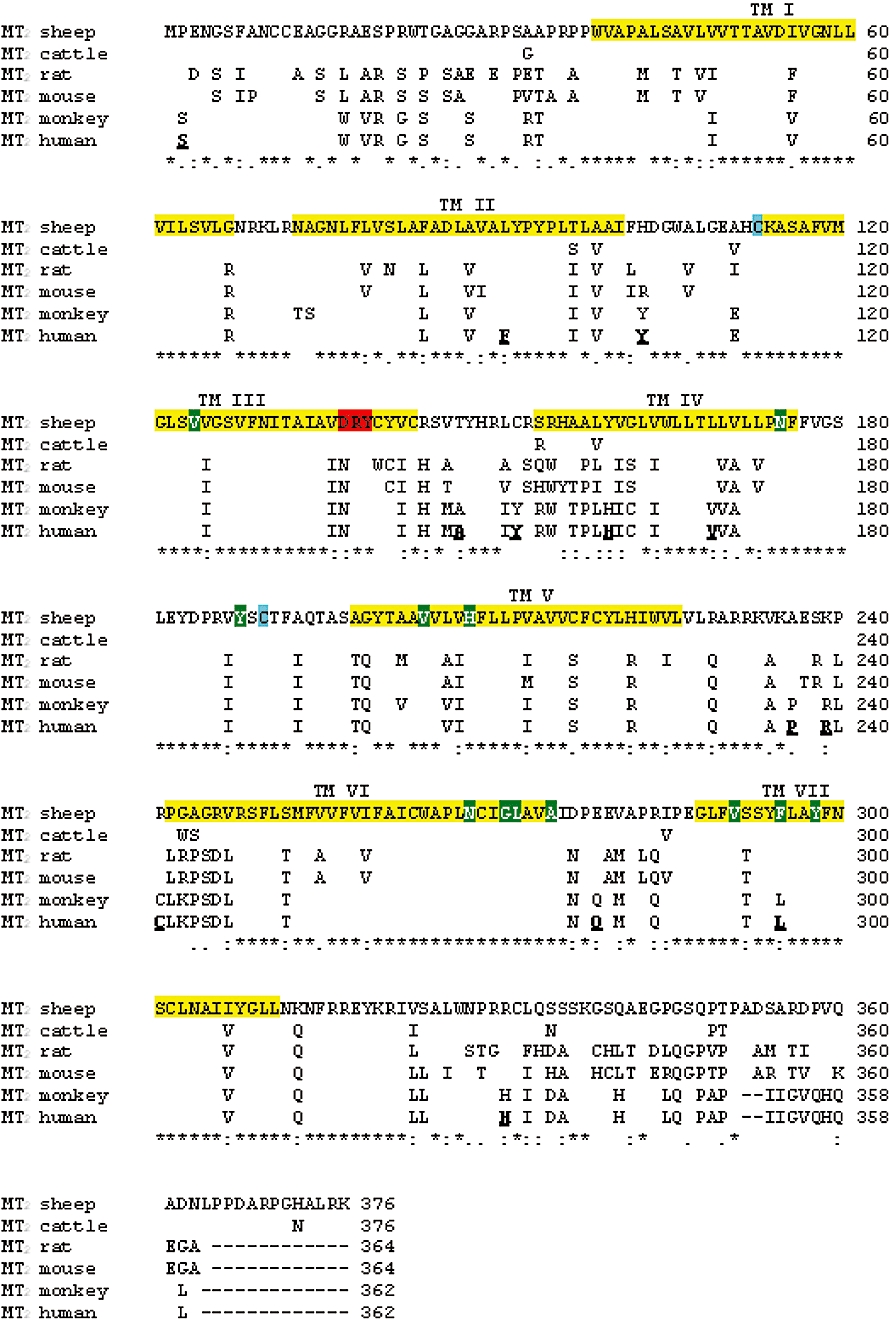
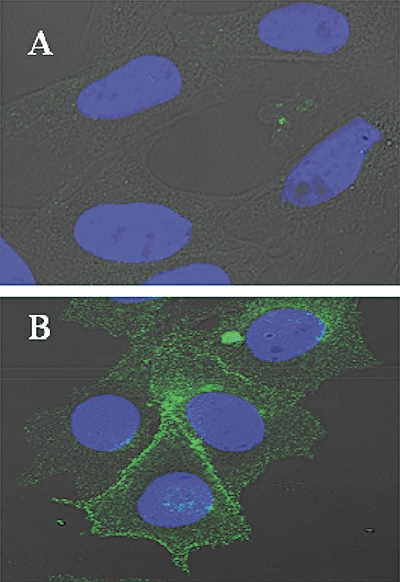
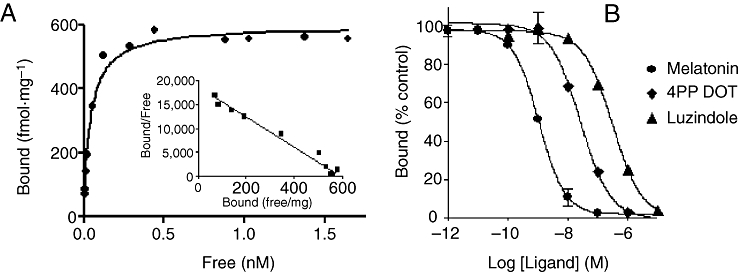
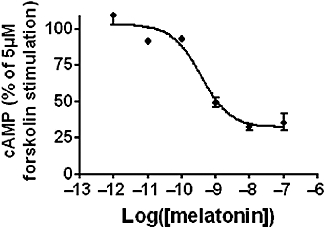
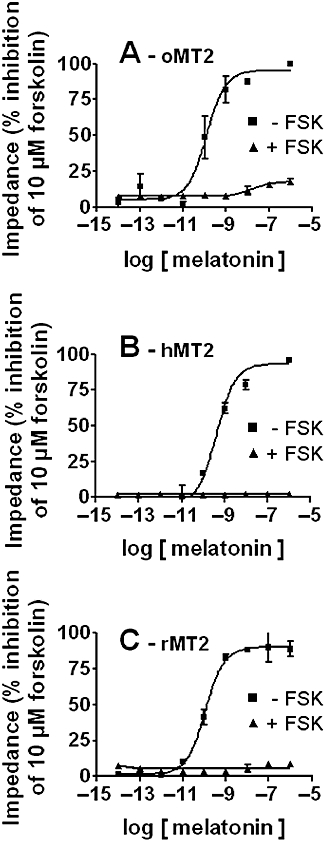
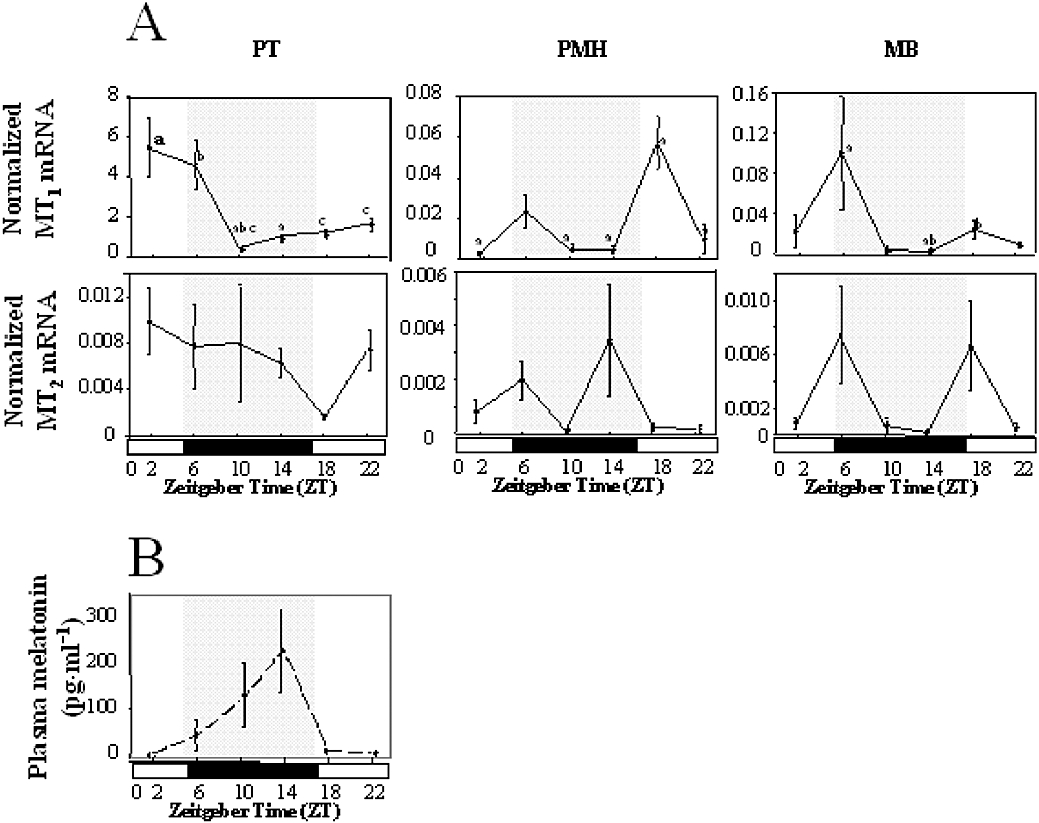
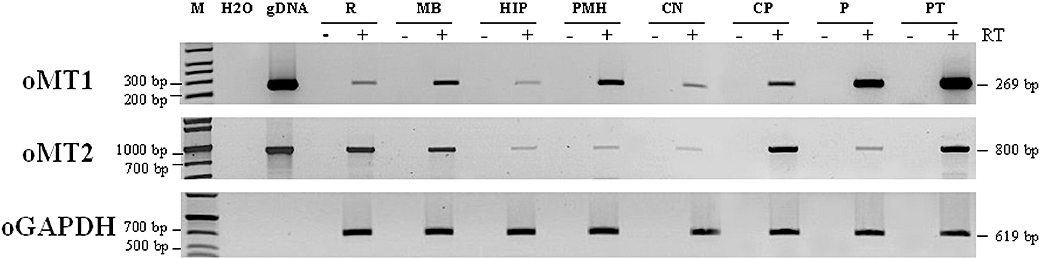
Key results: The ovine MT(2) receptor presents 96%, 72% and 67% identity with cattle, human and rat respectively. This MT(2) receptor stably expressed in CHO-K1 cells showed high-affinity 2[(125)I]-iodomelatonin binding (K(D)= 0.04 nM). The rank order of inhibition of 2[(125)I]-iodomelatonin binding by melatonin, 4-phenyl-2-propionamidotetralin and luzindole was similar to that exhibited by MT(2) receptors of other species (melatonin > 4-phenyl-2-propionamidotetralin > luzindole). However, its pharmacological profile was closer to that of rat, rather than human MT(2) receptors. Functionally, the ovine MT(2) receptors were coupled to G(i) proteins leading to inhibition of adenylyl cyclase, as the other melatonin receptors. In sheep brain, MT(2) mRNA was expressed in pars tuberalis, choroid plexus and retina, and moderately in mammillary bodies. Real-time polymerase chain reaction showed that in sheep pars tuberalis, premammillary hypothalamus and mammillary bodies, the temporal pattern of expression of MT(1) and MT(2) mRNA was not parallel in the three tissues.
Conclusion and implications: Co-expression of MT(1) and MT(2) receptors in all analysed sheep brain tissues suggests that MT(2) receptors may participate in melatonin regulation of seasonal anovulatory activity in ewes by modulating MT(1) receptor action.
Figures
References
-
- Alarma-Estrany P, Pintor J. Melatonin receptors in the eye: location, second messengers and role in ocular physiology. Pharmacol Ther. 2007;113:507–522. - PubMed
-
- Arendt J. Melatonin: characteristics, concerns, and prospects. J Biol Rhythms. 2005;20:291–303. - PubMed
-
- Audinot V, Mailliet F, Lahaye C, Bonnaud A, Le Gall A, Amosse A, et al. New selective ligands of human cloned melatonin MT1 and MT2 receptors. Naunyn Schmiedebergs Arch Pharmacol. 2003;367:553–561. - PubMed
-
- Audinot V, Bonnaud A, Grandcolas L, Rodriguez M, Nagel N, Galizzi JP, et al. Molecular cloning and binding characteristics of rat melatonin MT1 and MT2 receptors. Biochem Pharmacol. 2008;75:2007–2015. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
