

Protection of epithelial barrier function by the Crohn's disease associated gene protein tyrosine phosphatase n2
- PMID: 19818778
- PMCID: PMC2855721
- DOI: 10.1053/j.gastro.2009.07.078
Protection of epithelial barrier function by the Crohn's disease associated gene protein tyrosine phosphatase n2
Abstract
Background & aims: Protein tyrosine phosphatase N2 (PTPN2) has been identified as a Crohn's disease (CD) candidate gene. However, a role for PTPN2 in the pathogenesis of CD has not been identified. Increased permeability of the intestinal epithelium is believed to contribute prominently to CD. The aim of this study was to determine a possible role for PTPN2 in CD pathogenesis.
Methods: Intestinal epithelial cell (IEC) lines T(84) and HT29cl.19a were used in all studies. Protein analysis was performed by Western blotting, and protein knockdown was induced by small interfering RNA. Primary samples were from control and CD patients.
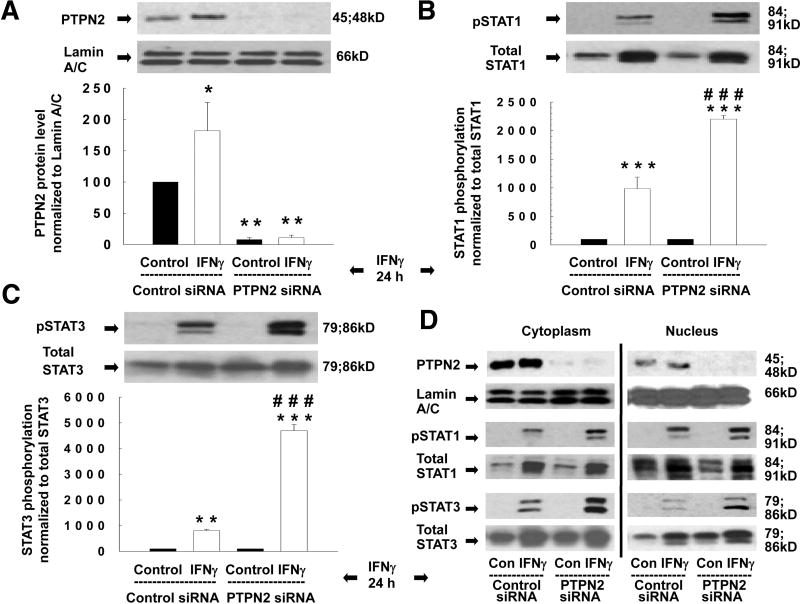
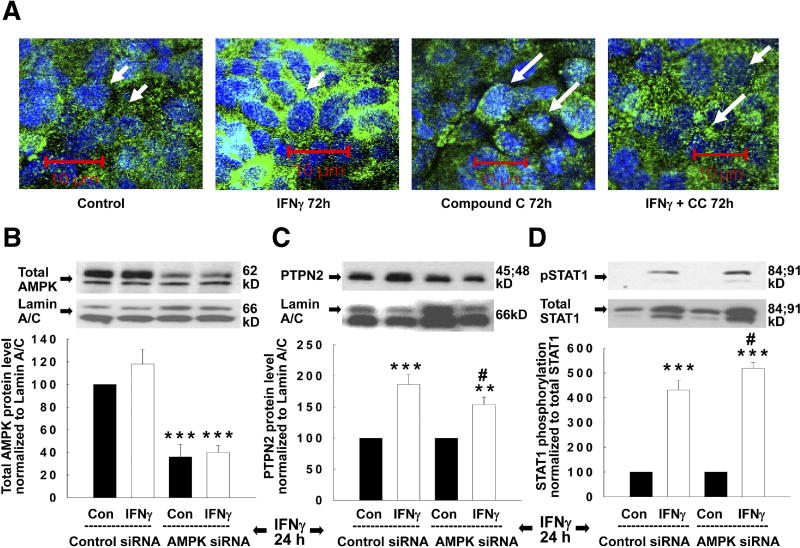
Results: Here, we demonstrate increased PTPN2 expression in CD intestinal biopsy specimens and that the proinflammatory cytokine interferon (IFN)-gamma increases PTPN2 expression and activity in IEC. Moreover, IFN-gamma-induced STAT1 and STAT3 phosphorylation in IEC is enhanced by PTPN2 knockdown. The cellular energy sensor adenosine monophosphate-activated protein kinase partially regulates the IFN-gamma-induced effects on PTPN2. Additionally, PTPN2 knockdown potentiates IFN-gamma-induced increases in epithelial permeability, accompanied by elevated expression of the pore-forming protein claudin-2.
Conclusions: PTPN2 is activated by IFN-gamma and limits IFN-gamma-induced signalling and consequent barrier defects. These data suggest a functional role for PTPN2 in maintaining the intestinal epithelial barrier and in the pathophysiology of CD.
Conflict of interest statement
The authors declare that they have no competing financial interests.
Figures




References
-
- Tonks NK, Neel BG. Combinatorial control of the specificity of protein tyrosine phosphatases. Curr Opin Cell Biol. 2001;13:182–195. - PubMed
-
- Franke A, Balschun T, Karlsen TH, Hedderich J, May S, Lu T, Schuldt D, Nikolaus S, Rosenstiel P, Krawczak M, Schreiber S. Replication of signals from recent studies of Crohn’s disease identifies previously unknown disease loci for ulcerative colitis. Nat Genet. 2008 - PubMed
-
- Parkes M, Barrett JC, Prescott NJ, Tremelling M, Anderson CA, Fisher SA, Roberts RG, Nimmo ER, Cummings FR, Soars D, Drummond H, Lees CW, Khawaja SA, Bagnall R, Burke DA, Todhunter CE, Ahmad T, Onnie CM, McArdle W, Strachan D, Bethel G, Bryan C, Lewis CM, Deloukas P, Forbes A, Sanderson J, Jewell DP, Satsangi J, Mansfield JC, Cardon L, Mathew CG. Sequence variants in the autophagy gene IRGM and multiple other replicating loci contribute to Crohn’s disease susceptibility. Nat Genet. 2007;39:830–832. - PMC - PubMed
-
- Fisher SA, Tremelling M, Anderson CA, Gwilliam R, Bumpstead S, Prescott NJ, Nimmo ER, Massey D, Berzuini C, Johnson C, Barrett JC, Cummings FR, Drummond H, Lees CW, Onnie CM, Hanson CE, Blaszczyk K, Inouye M, Ewels P, Ravindrarajah R, Keniry A, Hunt S, Carter M, Watkins N, Ouwehand W, Lewis CM, Cardon L, Lobo A, Forbes A, Sanderson J, Jewell DP, Mansfield JC, Deloukas P, Mathew CG, Parkes M, Satsangi J. Genetic determinants of ulcerative colitis include the ECM1 locus and five loci implicated in Crohn’s disease. Nat Genet. 2008 - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
