

Convergence of 3',5'-cyclic adenosine 5'-monophosphate/protein kinase A and glycogen synthase kinase-3beta/beta-catenin signaling in corpus luteum progesterone synthesis
- PMID: 19819952
- PMCID: PMC3213761
- DOI: 10.1210/en.2009-0771
Convergence of 3',5'-cyclic adenosine 5'-monophosphate/protein kinase A and glycogen synthase kinase-3beta/beta-catenin signaling in corpus luteum progesterone synthesis
Abstract
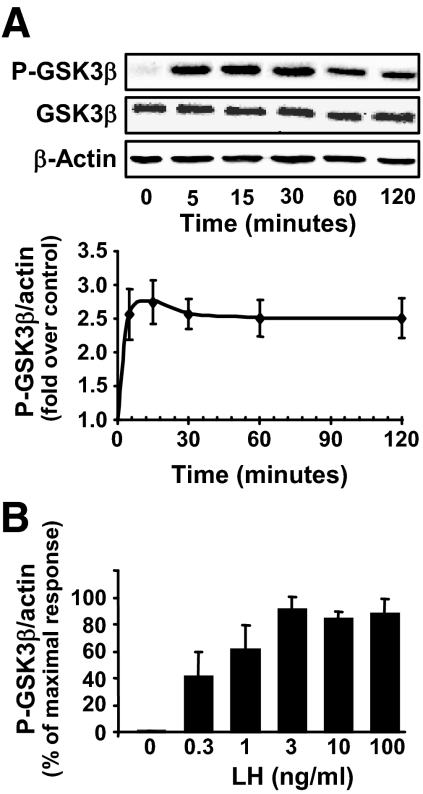
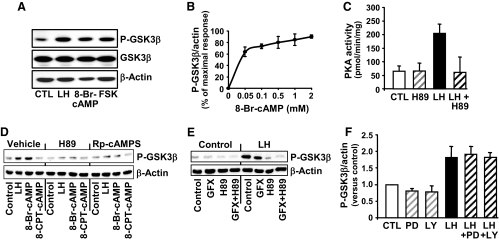
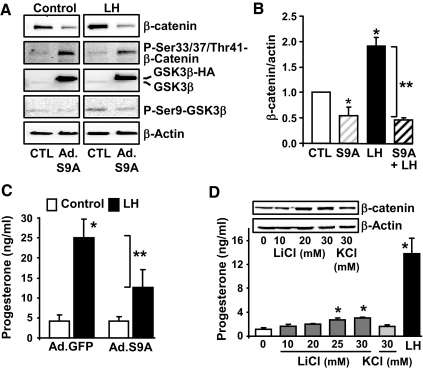
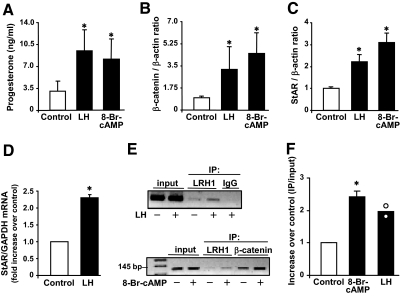
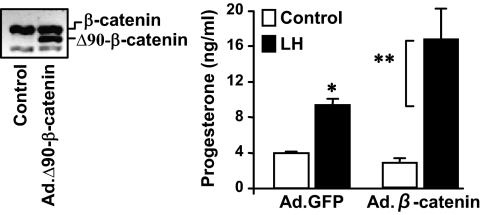
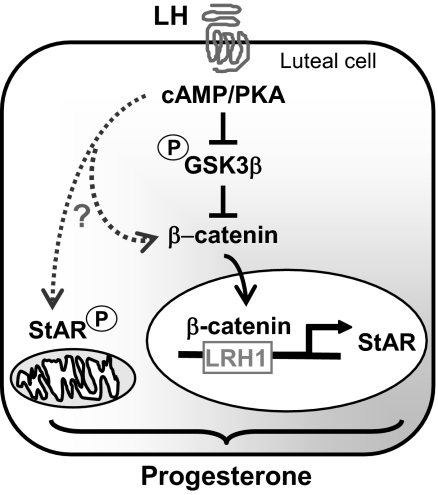
Progesterone secretion by the steroidogenic cells of the corpus luteum (CL) is essential for reproduction. Progesterone synthesis is under the control of LH, but the exact mechanism of this regulation is unknown. It is established that LH stimulates the LH receptor/choriogonadotropin receptor, a G-protein coupled receptor, to increase cAMP and activate cAMP-dependent protein kinase A (PKA). In the present study, we tested the hypothesis that cAMP/PKA-dependent regulation of the Wnt pathway components glycogen synthase kinase (GSK)-3beta and beta-catenin contributes to LH-dependent steroidogenesis in luteal cells. We observed that LH via a cAMP/PKA-dependent mechanism stimulated the phosphorylation of GSK3beta at N-terminal Ser9 causing its inactivation and resulted in the accumulation of beta-catenin. Overexpression of N-terminal truncated beta-catenin (Delta90 beta-catenin), which lacks the phosphorylation sites responsible for its destruction, significantly augmented LH-stimulated progesterone secretion. In contrast, overexpression of a constitutively active mutant of GSK3beta (GSK-S9A) reduced beta-catenin levels and inhibited LH-stimulated steroidogenesis. Chromatin immunoprecipitation assays demonstrated the association of beta-catenin with the proximal promoter of the StAR gene, a gene that expresses the steroidogenic acute regulatory protein, which is a cholesterol transport protein that controls a rate-limiting step in steroidogenesis. Collectively these data suggest that cAMP/PKA regulation of GSK3beta/beta-catenin signaling may contribute to the acute increase in progesterone production in response to LH.
Figures
References
-
- Davis JS, Rueda BR 2002 The corpus luteum: an ovarian structure with maternal instincts and suicidal tendencies. Front Biosci 7:d1949–d1978 - PubMed
-
- Stouffer RL 2003 Progesterone as a mediator of gonadotrophin action in the corpus luteum: beyond steroidogenesis. Hum Reprod Update 9:99–117 - PubMed
-
- Narko K, Saukkonen K, Ketola I, Bützow R, Heikinheimo M, Ristimäki A 2001 Regulated expression of prostaglandin E(2) receptors EP2 and EP4 in human ovarian granulosa-luteal cells. J Clin Endocrinol Metab 86:1765–1768 - PubMed
-
- Regan JW 2003 EP2 and EP4 prostanoid receptor signaling. Life Sci 74:143–153 - PubMed
-
- Niswender GD 2002 Molecular control of luteal secretion of progesterone. Reproduction 123:333–339 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
