

Phenotypic plasticity of adventitious rooting in Arabidopsis is controlled by complex regulation of AUXIN RESPONSE FACTOR transcripts and microRNA abundance
- PMID: 19820192
- PMCID: PMC2782293
- DOI: 10.1105/tpc.108.064758
Phenotypic plasticity of adventitious rooting in Arabidopsis is controlled by complex regulation of AUXIN RESPONSE FACTOR transcripts and microRNA abundance
Abstract
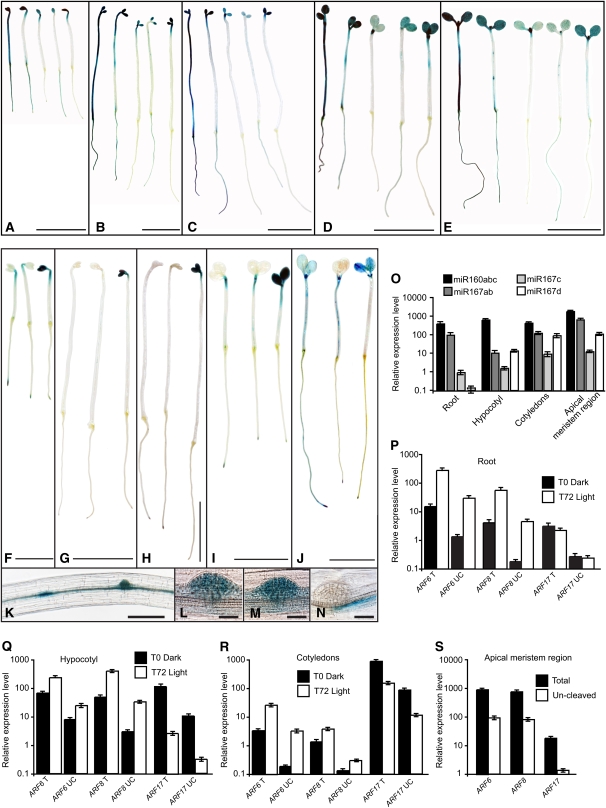
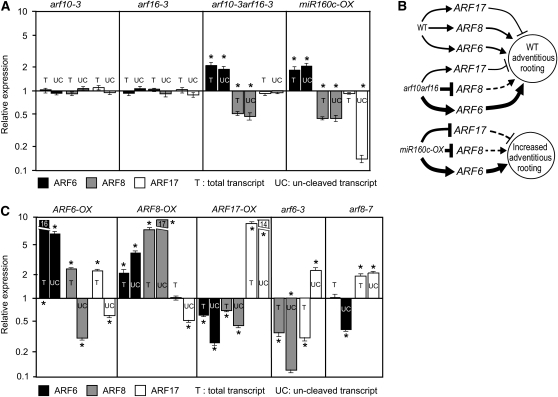
The development of shoot-borne roots, or adventitious roots, is indispensable for mass propagation of elite genotypes. It is a complex genetic trait with a high phenotypic plasticity due to multiple endogenous and environmental regulatory factors. We demonstrate here that a subtle balance of activator and repressor AUXIN RESPONSE FACTOR (ARF) transcripts controls adventitious root initiation. Moreover, microRNA activity appears to be required for fine-tuning of this process. Thus, ARF17, a target of miR160, is a negative regulator, and ARF6 and ARF8, targets of miR167, are positive regulators of adventitious rooting. The three ARFs display overlapping expression domains, interact genetically, and regulate each other's expression at both transcriptional and posttranscriptional levels by modulating miR160 and miR167 availability. This complex regulatory network includes an unexpected feedback regulation of microRNA homeostasis by direct and nondirect target transcription factors. These results provide evidence of microRNA control of phenotypic variability and are a significant step forward in understanding the molecular mechanisms regulating adventitious rooting.
Figures




References
-
- Allen, E., Xie, Z., Gustafson, A.M., and Carrington, J.C. (2005). MicroRNA-directed phasing during trans-acting siRNA biogenesis in plants. Cell 121 207–221. - PubMed
-
- Clough, S.J., and Bent, A.F. (1998). Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 16 735–743. - PubMed
-
- Franco-Zorrilla, J.M., Valli, A., Todesco, M., Mateos, I., Puga, M.I., Rubio-Somoza, I., Leyva, A., Weigel, D., Garcia, J.A., and Paz-Ares, J. (2007). Target mimicry provides a new mechanism for regulation of microRNA activity. Nat. Genet. 39 1033–1037. - PubMed
-
- Gasciolli, V., Mallory, A.C., Bartel, D.P., and Vaucheret, H. (2005). Partially redundant functions of Arabidopsis DICER-like enzymes and a role for DCL4 in producing trans-acting siRNAs. Curr. Biol. 15 1494–1500. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
