

Carbon and nitrogen nutrient balance signaling in plants
- PMID: 19820356
- PMCID: PMC2710548
- DOI: 10.4161/psb.4.7.8540
Carbon and nitrogen nutrient balance signaling in plants
Abstract
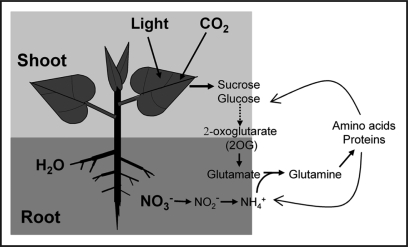
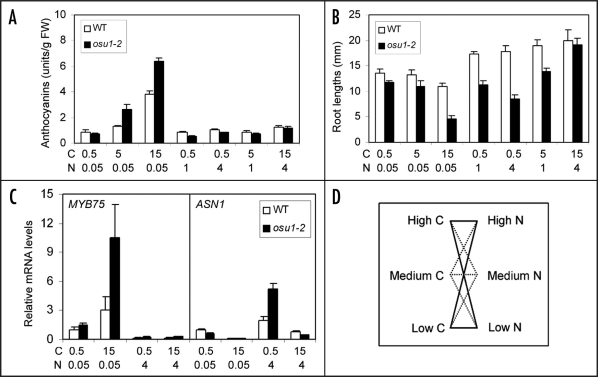
Cellular carbon (C) and nitrogen (N) metabolism must be tightly coordinated to sustain optimal growth and development for plants and other cellular organisms. Furthermore, C/N balance is also critical for the ecosystem response to elevated atmospheric CO(2). Despite numerous physiological and molecular studies in C/N balance or ratio response, very few genes have been shown to play important roles in C/N balance signaling. During recent five years, exciting progress was made through genetic and genomic studies. Several DNA microarray studies have shown that more than half of the transcriptome is regulated by C, N and the C-N combination. Three genetic studies involving distinct bioassays have demonstrated that a putative nitrate transporter (NTR2.1), a putative glutamate receptor (GLR1.1) and a putative methyltransferase (OSU1) have important functions in the C/N balance response. OSU1 is identical to QUA2/TSD2 which has been implicated to act in cell wall biogenesis, indicating a link between cell wall property and the C/N balance signaling. Given that many investigations are only focused on C alone or N alone, the C/N balance bioassays and gene expression patterns are discussed to assist phenotypic characterization of C/N balance signaling. Further, re-examination of those previously reported sugar or nitrogen responsive genes in C/N balance response may be necessary to dissect the C/N signaling pathways. In addition, key components involved in C-N interactions in bacterial, yeast and animal systems and whether they are functionally conserved in plants are discussed. These rapid advances have provided the first important step towards the construction of the complex yet elegant C/N balance signaling networks in plants.
Figures

References
-
- Coruzzi GM, Zhou L. Carbon and nitrogen sensing and signaling in plants: emerging ‘matrix effects’. Curr Opin Plant Biol. 2001;4:247–253. - PubMed
-
- Schofield RA, Bi YM, Kant S, Rothstein SJ. Overexpression of STP13, a hexose transporter, improves plant growth and nitrogen use in Arabidopsis thaliana seedlings. Plant Cell Environ. 2009;32:271–285. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources