

Segregation of short-wavelength-sensitive (S) cone signals in the macaque dorsal lateral geniculate nucleus
- PMID: 19821840
- PMCID: PMC2777259
- DOI: 10.1111/j.1460-9568.2009.06939.x
Segregation of short-wavelength-sensitive (S) cone signals in the macaque dorsal lateral geniculate nucleus
Abstract
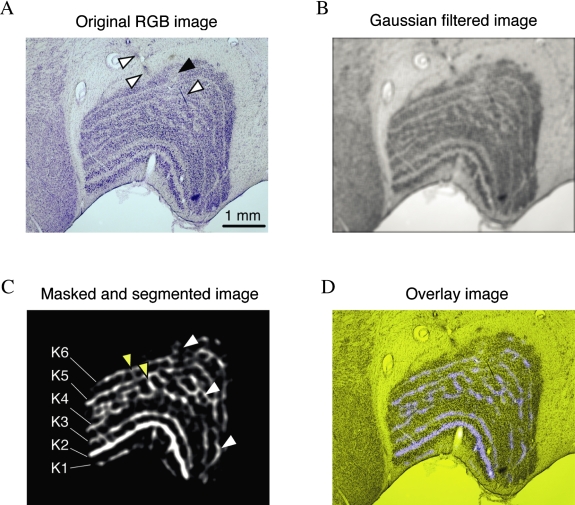
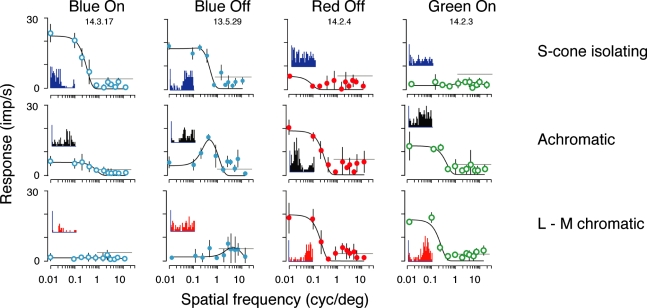
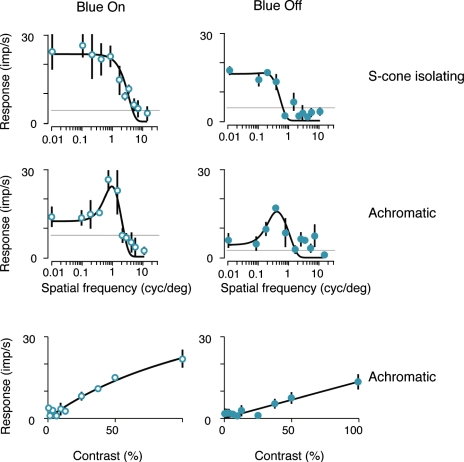
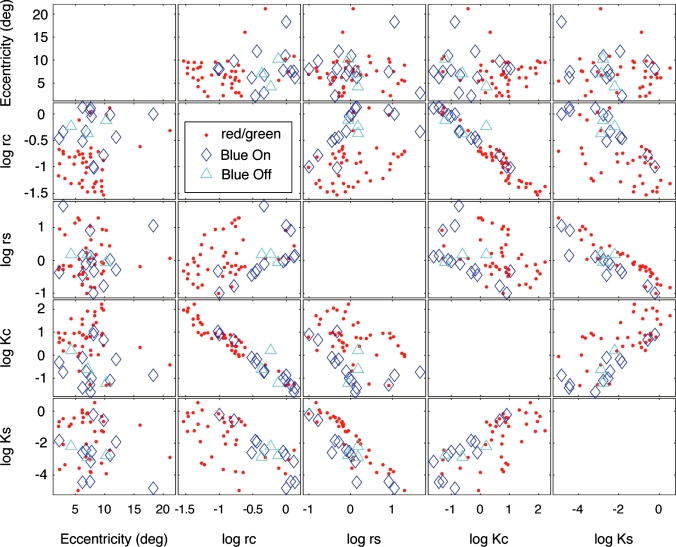
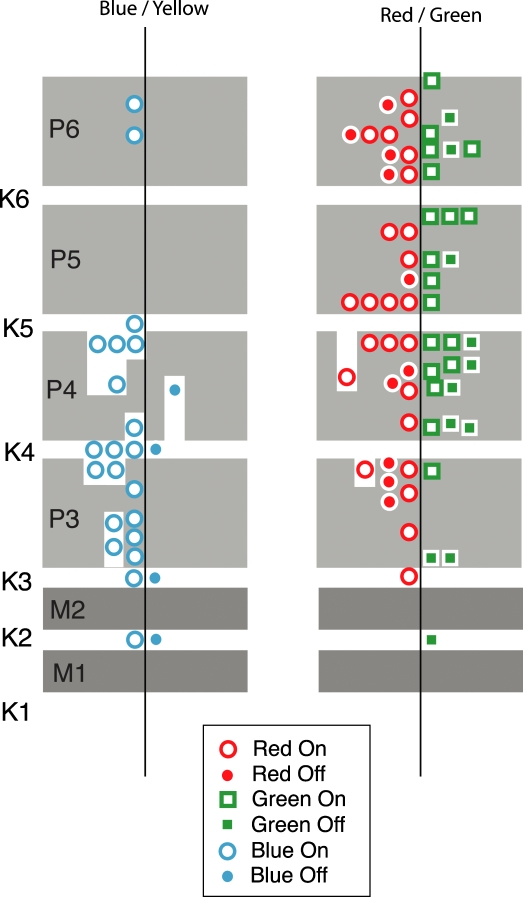
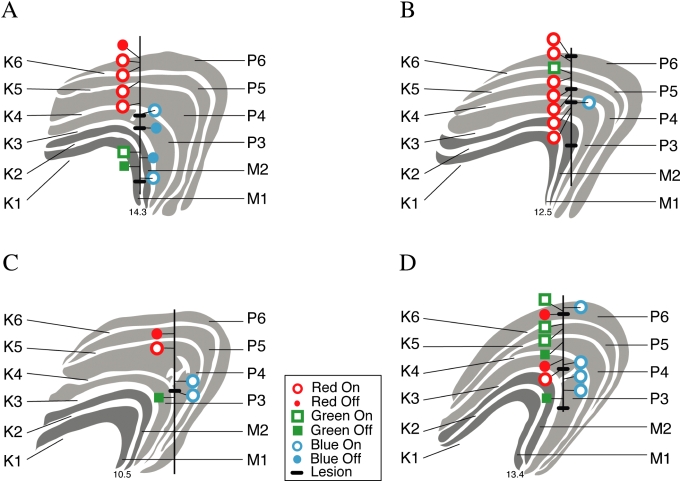
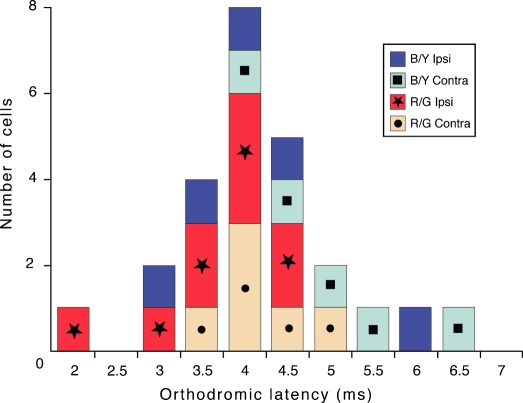
An important problem in the study of the mammalian visual system is whether functionally different retinal ganglion cell types are anatomically segregated further up along the central visual pathway. It was previously demonstrated that, in a New World diurnal monkey (marmoset), the neurones carrying signals from the short-wavelength-sensitive (S) cones [blue-yellow (B/Y)-opponent cells] are predominantly located in the koniocellular layers of the dorsal lateral geniculate nucleus (LGN), whereas the red-green (R/G)-opponent cells carrying signals from the medium- and long-wavelength-sensitive cones are segregated in the parvocellular layers. Here, we used extracellular single-unit recordings followed by histological reconstruction to investigate the distribution of color-selective cells in the LGN of the macaque, an Old World diurnal monkey. Cells were classified using cone-isolating stimuli to identify their cone inputs. Our results indicate that the majority of cells carrying signals from S-cones are located either in the koniocellular layers or in the 'koniocellular bridges' that fully or partially span the parvocellular layers. By contrast, the R/G-opponent cells are located in the parvocellular layers. We conclude that anatomical segregation of B/Y- and R/G-opponent afferent signals for color vision is common to the LGNs of New World and Old World diurnal monkeys.
Figures
References
-
- Ahmad A, Spear PD. Effects of aging on the size, density, and number of rhesus monkey lateral geniculate neurons. J. Comp. Neurol. 1993;334:631–643. - PubMed
-
- Brainard DH. The Psychophysics Toolbox. Spat. Vis. 1997;10:443–446. - PubMed
-
- Casagrande VA, Royal DW, Sáry G. Extraretinal inputs and feedback mechanisms to the lateral geniculate nucleus (LGN) In: Kremers J, editor. The Primate Visual System: A Comparative Approach. Hoboken, New Jersey: John Wiley & Sons; 2005. pp. 191–211.
-
- Chatterjee S, Callaway EM. Parallel colour-opponent pathways to primary visual cortex. Nature. 2003;426:668–671. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
