

ATP released by electrical stimuli elicits calcium transients and gene expression in skeletal muscle
- PMID: 19822518
- PMCID: PMC2787310
- DOI: 10.1074/jbc.M109.057315
ATP released by electrical stimuli elicits calcium transients and gene expression in skeletal muscle
Abstract
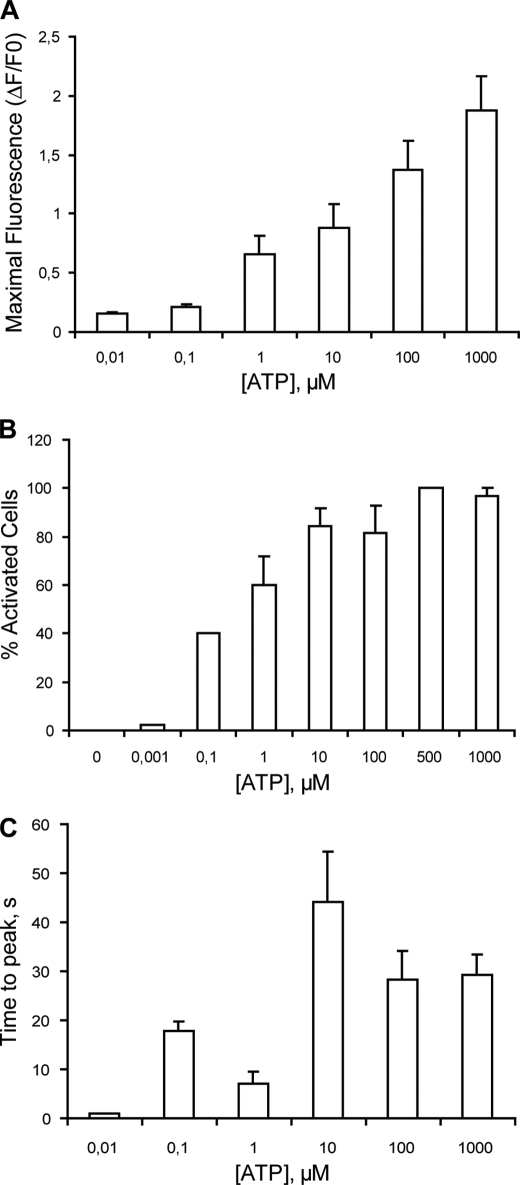
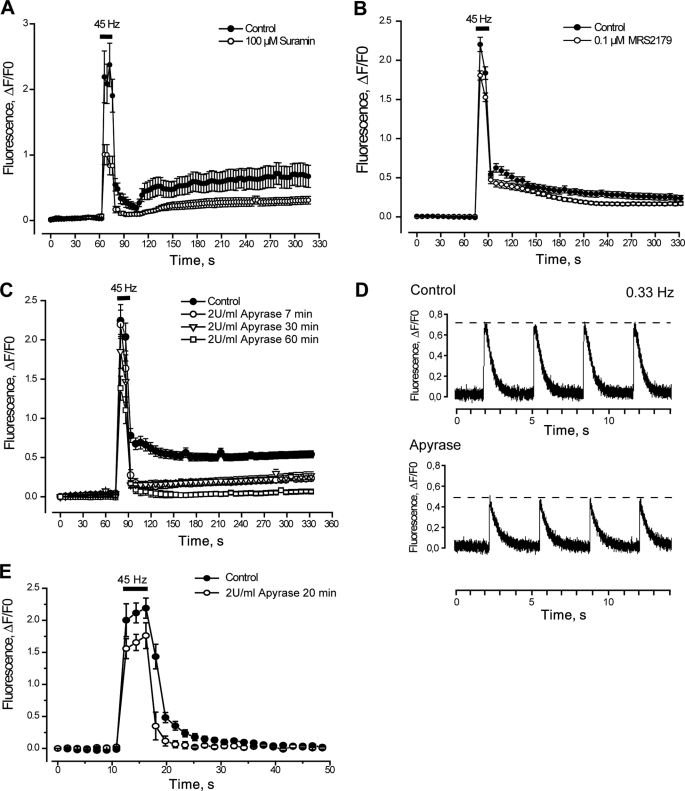
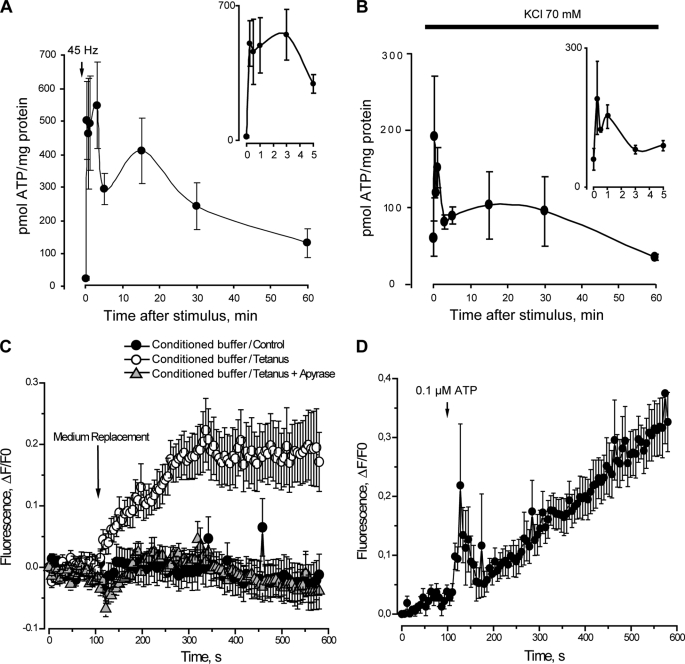
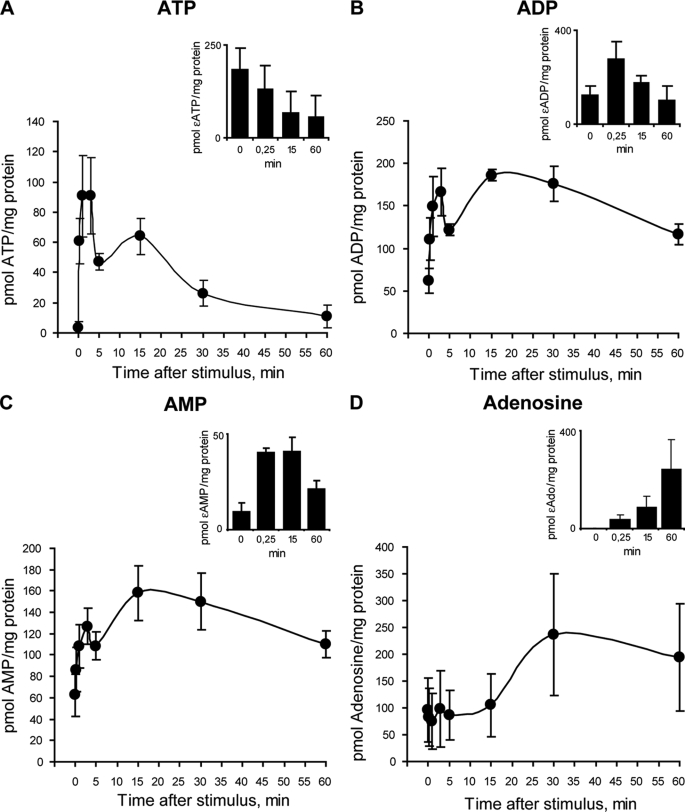
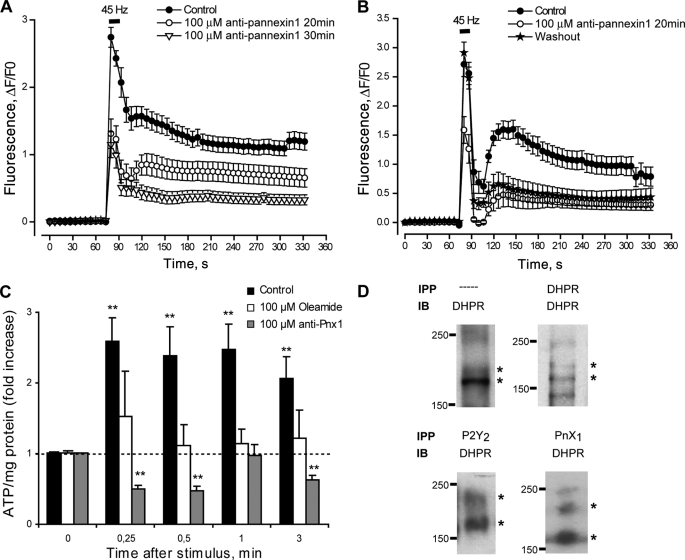
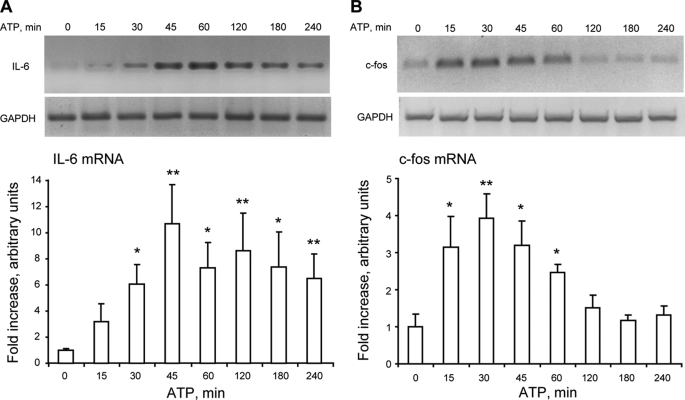
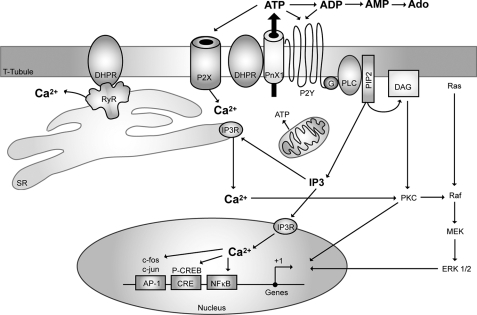
ATP released from cells is known to activate plasma membrane P2X (ionotropic) or P2Y (metabotropic) receptors. In skeletal muscle cells, depolarizing stimuli induce both a fast calcium signal associated with contraction and a slow signal that regulates gene expression. Here we show that nucleotides released to the extracellular medium by electrical stimulation are partly involved in the fast component and are largely responsible for the slow signals. In rat skeletal myotubes, a tetanic stimulus (45 Hz, 400 1-ms pulses) rapidly increased extracellular levels of ATP, ADP, and AMP after 15 s to 3 min. Exogenous ATP induced an increase in intracellular free Ca(2+) concentration, with an EC(50) value of 7.8 +/- 3.1 microm. Exogenous ADP, UTP, and UDP also promoted calcium transients. Both fast and slow calcium signals evoked by tetanic stimulation were inhibited by either 100 mum suramin or 2 units/ml apyrase. Apyrase also reduced fast and slow calcium signals evoked by tetanus (45 Hz, 400 0.3-ms pulses) in isolated mouse adult skeletal fibers. A likely candidate for the ATP release pathway is the pannexin-1 hemichannel; its blockers inhibited both calcium transients and ATP release. The dihydropyridine receptor co-precipitated with both the P2Y(2) receptor and pannexin-1. As reported previously for electrical stimulation, 500 mum ATP significantly increased mRNA expression for both c-fos and interleukin 6. Our results suggest that nucleotides released during skeletal muscle activity through pannexin-1 hemichannels act through P2X and P2Y receptors to modulate both Ca(2+) homeostasis and muscle physiology.
Figures
References
-
- Grabner M., Dirksen R. T., Suda N., Beam K. G. (1999) J. Biol. Chem. 274, 21913–21919 - PubMed
-
- Lamb G. D. (2002) Front. Biosci. 7, d834–d842 - PubMed
-
- Tanabe T., Beam K. G., Adams B. A., Niidome T., Numa S. (1990) Nature 346, 567–569 - PubMed
-
- Carrasco M. A., Riveros N., Ríos J., Müller M., Torres F., Pineda J., Lantadilla S., Jaimovich E. (2003) Am. J. Physiol. Cell Physiol. 284, C1438–C1447 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
