

Evidence for the Existence in Arabidopsis thaliana of the Proteasome Proteolytic Pathway: ACTIVATION IN RESPONSE TO CADMIUM
- PMID: 19822524
- PMCID: PMC2790970
- DOI: 10.1074/jbc.M109.035394
Evidence for the Existence in Arabidopsis thaliana of the Proteasome Proteolytic Pathway: ACTIVATION IN RESPONSE TO CADMIUM
Abstract
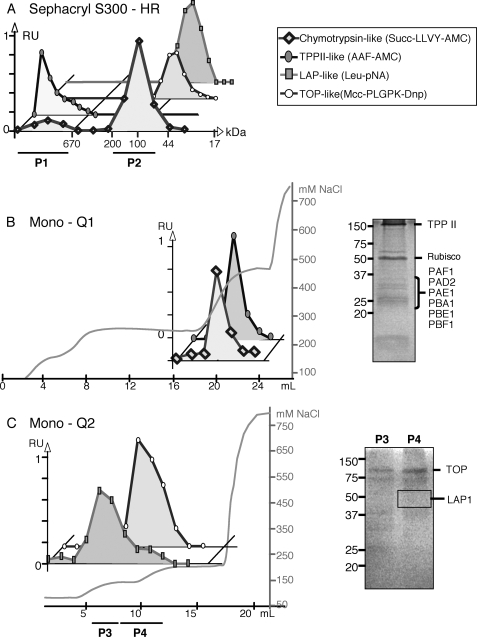
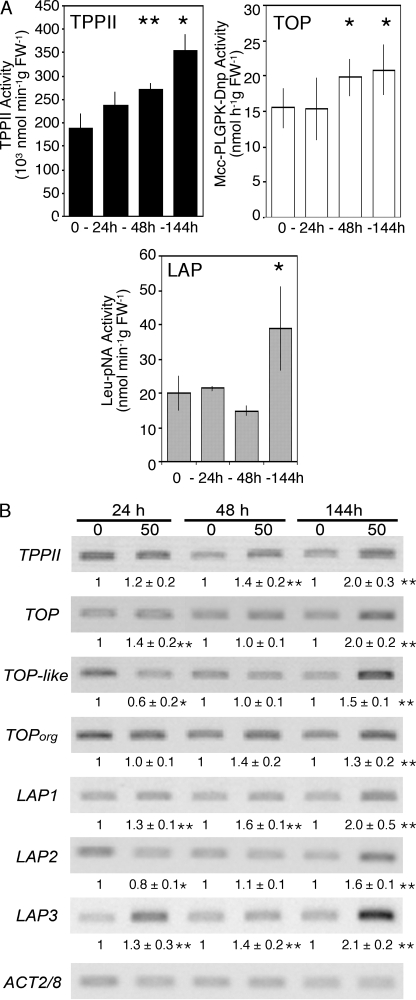
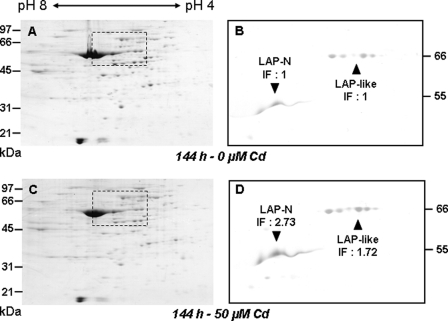
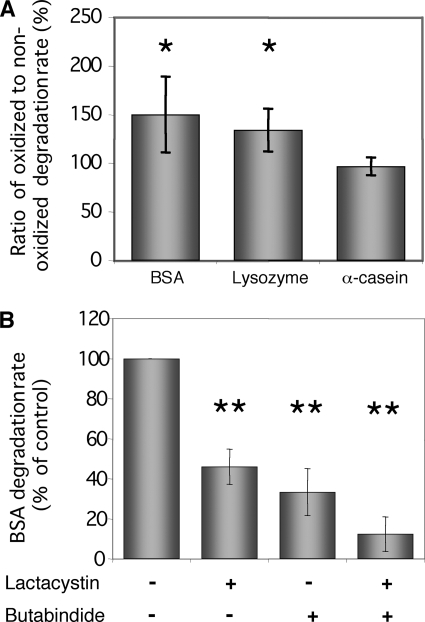
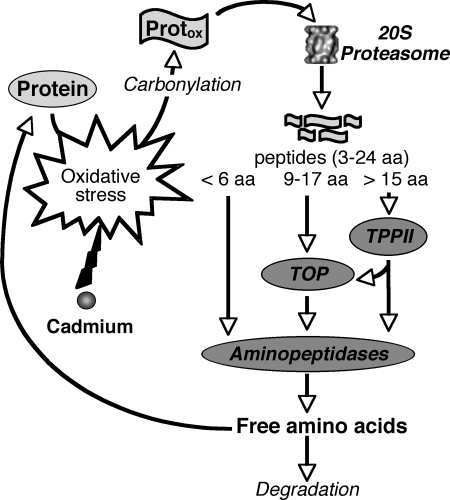
Heavy metals are known to generate reactive oxygen species that lead to the oxidation and fragmentation of proteins, which become toxic when accumulated in the cell. In this study, we investigated the role of the proteasome during cadmium stress in the leaves of Arabidopsis thaliana plants. Using biochemical and proteomics approaches, we present the first evidence of an active proteasome pathway in plants. We identified and characterized the peptidases acting sequentially downstream from the proteasome in animal cells as follows: tripeptidyl-peptidase II, thimet oligopeptidase, and leucine aminopeptidase. We investigated the proteasome proteolytic pathway response in the leaves of 6-week-old A. thaliana plants grown hydroponically for 24, 48, and 144 h in the presence or absence of 50 mum cadmium. The gene expression and proteolytic activity of the proteasome and the different proteases of the pathway were found to be up-regulated in response to cadmium. In an in vitro assay, oxidized bovine serum albumin and lysozyme were more readily degraded in the presence of 20 S proteasome and tripeptidyl-peptidase II than their nonoxidized form, suggesting that oxidized proteins are preferentially degraded by the Arabidopsis 20 S proteasome pathway. These results show that, in response to cadmium, the 20 S proteasome proteolytic pathway is up-regulated at both RNA and activity levels in Arabidopsis leaves and may play a role in degrading oxidized proteins generated by the stress.
Figures

References
-
- Sanita di Toppi L., Gabbrielli R. (1999) Environ. Exp. Bot. 41, 105–130
-
- Wagner G. (1993) Adv. Agron. 51, 173–212
-
- Lafuente A., Cabaleiro T., Caride A., Romero A. (2008) Electron. J. Environ. Agric. Food Chem. 7, 3363–3371
-
- Davies K. J. (2001) Biochimie 83, 301–310 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
