

Regulatory and pro-inflammatory phenotypes of myelin basic protein-autoreactive T cells in multiple sclerosis
- PMID: 19822525
- PMCID: PMC2779831
- DOI: 10.1093/intimm/dxp100
Regulatory and pro-inflammatory phenotypes of myelin basic protein-autoreactive T cells in multiple sclerosis
Abstract
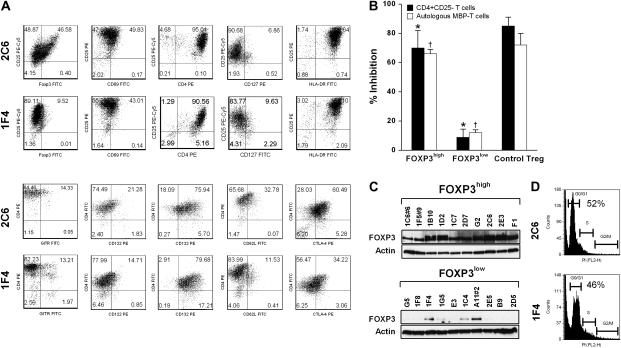
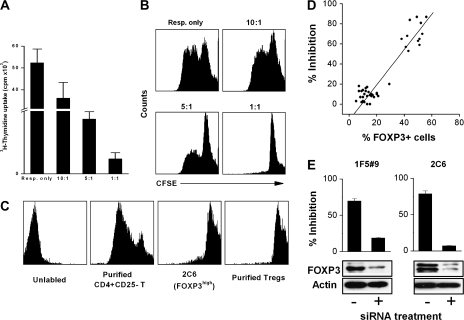
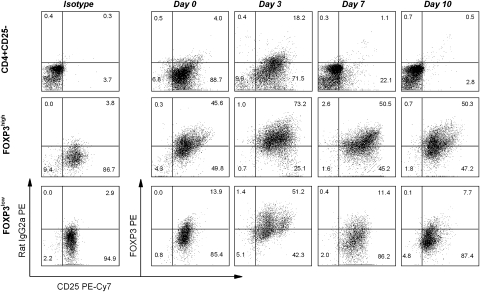
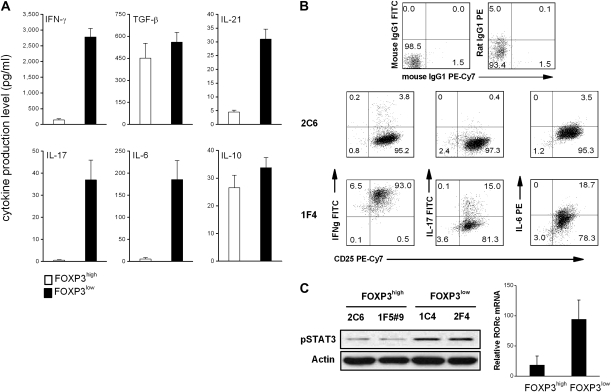
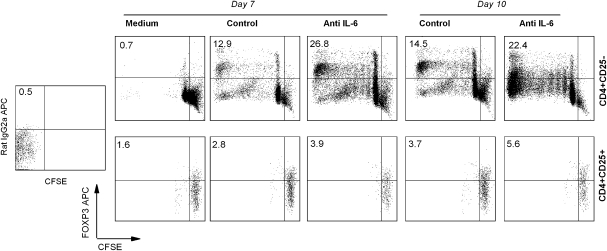
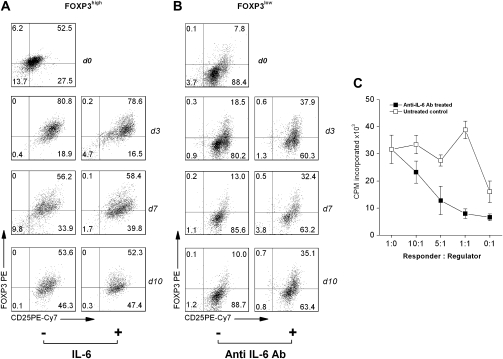
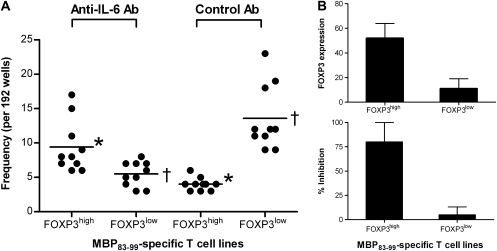
MBP-specific autoreactive T cells are considered pro-inflammatory T cells and thought to play an important role in the pathogenesis of multiple sclerosis (MS). Here, we report that MBP(83-99)-specific T cells generated from MS patients (n = 7) were comprised of pro-inflammatory and regulatory subsets of distinct phenotypes. The pro-inflammatory phenotype was characterized by high production of IFN-gamma, IL-6, IL-21 and IL-17 and low expression of FOXP3, whereas the regulatory subset expressed high levels of FOXP3 and exhibited potent regulatory functions. The regulatory subset of MBP-specific T cells appeared to expand from the CD4(+)CD25(-) T-cell pool. Their FOXP3 expression was stable, independent of the activation state and it correlated with suppressive function and inversely with the production of IFN-gamma, IL-6, IL-21 and IL-17. In contrast, the phenotype and function of FOXP3(low) MBP-specific T cells were adaptive and dependent on IL-6. The higher frequency of FOXP3(high) MBP-specific T cells was observed when IL-6 was neutralized in the culture of PBMC with MBP. The study provides new evidence that MBP-specific T cells are susceptible to pro-inflammatory cytokine milieu and act as either pro-inflammatory or regulatory T cells.
Figures
Similar articles
-
Cytokine phenotype of human autoreactive T cell clones specific for the immunodominant myelin basic protein peptide (83-99).J Neurosci Res. 1996 Sep 15;45(6):852-62. doi: 10.1002/(SICI)1097-4547(19960915)45:6<852::AID-JNR22>3.0.CO;2-W. J Neurosci Res. 1996. PMID: 8892097
-
Cytokine secretion profile of myelin basic protein-specific T cells in multiple sclerosis.Mult Scler. 2000 Apr;6(2):69-77. doi: 10.1177/135245850000600203. Mult Scler. 2000. PMID: 10773850
-
Cytokine secretion of myelin basic protein reactive T cells in patients with multiple sclerosis.J Neuroimmunol. 1998 Nov 2;91(1-2):1-9. doi: 10.1016/s0165-5728(98)00086-1. J Neuroimmunol. 1998. PMID: 9846813
-
Autoimmune pathogenesis of multiple sclerosis: role of autoreactive T lymphocytes and new immunotherapeutic strategies.Crit Rev Immunol. 1997;17(1):33-75. doi: 10.1615/critrevimmunol.v17.i1.20. Crit Rev Immunol. 1997. PMID: 9034723 Review.
-
Myelin basic protein-reactive T cells in multiple sclerosis: pathologic relevance and therapeutic targeting.Cytotechnology. 1994;16(3):181-7. doi: 10.1007/BF00749906. Cytotechnology. 1994. PMID: 7537052 Review.
Cited by
-
IL-21 and IL-21 receptor expression in lymphocytes and neurons in multiple sclerosis brain.Am J Pathol. 2011 Feb;178(2):794-802. doi: 10.1016/j.ajpath.2010.10.043. Am J Pathol. 2011. PMID: 21281812 Free PMC article.
-
The Molecular Mechanisms of Vitamin A Deficiency in Multiple Sclerosis.J Mol Neurosci. 2016 Sep;60(1):82-90. doi: 10.1007/s12031-016-0781-0. Epub 2016 Jun 29. J Mol Neurosci. 2016. PMID: 27356515 Review.
-
Forskolin improves experimental autoimmune encephalomyelitis in mice probably by inhibiting the calcium and the IL-17-STEAP4 signaling pathway.Heliyon. 2024 Aug 10;10(16):e36063. doi: 10.1016/j.heliyon.2024.e36063. eCollection 2024 Aug 30. Heliyon. 2024. PMID: 39229522 Free PMC article.
-
Stroke induces a rapid adaptive autoimmune response to novel neuronal antigens.Discov Med. 2015 May;19(106):381-92. Discov Med. 2015. PMID: 26105701 Free PMC article.
-
Adipocytokine profile, cytokine levels and foxp3 expression in multiple sclerosis: a possible link to susceptibility and clinical course of disease.PLoS One. 2013 Oct 3;8(10):e76555. doi: 10.1371/journal.pone.0076555. eCollection 2013. PLoS One. 2013. PMID: 24098530 Free PMC article.
References
-
- Ben-Nun A, Cohen IR. Experimental autoimmune encephalomyelitis (EAE) mediated by T cell lines: process of selection of lines and characterization of the cells. J. Immunol. 1982;129:303. - PubMed
-
- Ota K, Matsui M, Milford EL, Mackin GA, Weiner HL, Hafler DA. T-cell recognition of an immunodominant myelin basic protein epitope in multiple sclerosis. Nature. 1990;346:183. - PubMed
-
- Zamvil S, Nelson P, Trotter J, et al. T-cell clones specific for myelin basic protein induce chronic relapsing paralysis and demyelination. Nature. 1985;317:355. - PubMed
-
- Mokhtarian F, McFarlin DE, Raine CS. Adoptive transfer of myelin basic protein-sensitized T cells produces chronic relapsing demyelinating disease in mice. Nature. 1984;309:356. - PubMed
-
- Zhang J, Markovic-Plese S, Lacet B, Raus J, Weiner HL, Hafler DA. Increased frequency of interleukin 2-responsive T cells specific for myelin basic protein and proteolipid protein in peripheral blood and cerebrospinal fluid of patients with multiple sclerosis. J. Exp. Med. 1994;179:973. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
