

Two fundamentally distinct PCNA interaction peptides contribute to chromatin assembly factor 1 function
- PMID: 19822659
- PMCID: PMC2786881
- DOI: 10.1128/MCB.01051-09
Two fundamentally distinct PCNA interaction peptides contribute to chromatin assembly factor 1 function
Abstract
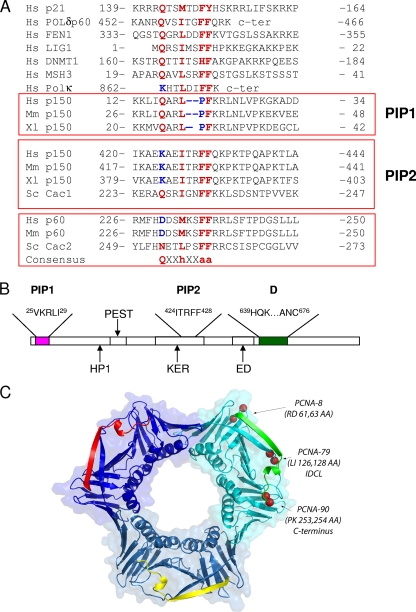
Chromatin assembly factor 1 (CAF-1) deposits histones H3 and H4 rapidly behind replication forks through an interaction with the proliferating cell nuclear antigen (PCNA), a DNA polymerase processivity factor that also binds to a number of replication enzymes and other proteins that act on nascent DNA. The mechanisms that enable CAF-1 and other PCNA-binding proteins to function harmoniously at the replication fork are poorly understood. Here we report that the large subunit of human CAF-1 (p150) contains two distinct PCNA interaction peptides (PIPs). The N-terminal PIP binds strongly to PCNA in vitro but, surprisingly, is dispensable for nucleosome assembly and only makes a modest contribution to targeting p150 to DNA replication foci in vivo. In contrast, the internal PIP (PIP2) lacks one of the highly conserved residues of canonical PIPs and binds weakly to PCNA. Surprisingly, PIP2 is essential for nucleosome assembly during DNA replication in vitro and plays a major role in targeting p150 to sites of DNA replication. Unlike canonical PIPs, such as that of p21, the two p150 PIPs are capable of preferentially inhibiting nucleosome assembly, rather than DNA synthesis, suggesting that intrinsic features of these peptides are part of the mechanism that enables CAF-1 to function behind replication forks without interfering with other PCNA-mediated processes.
Figures







References
-
- Bienko, M., C. M. Green, N. Crosetto, F. Rudolf, G. Zapart, B. Coull, P. Kannouche, G. Wider, M. Peter, A. R. Lehmann, K. Hofmann, and I. Dikic. 2005. Ubiquiting-binding domains in Y-family polymerases regulate translesion synthesis. Science 310:1821-1824. - PubMed
-
- Bowman, G. D., M. O'Donnell, and J. Kuriyan. 2004. Structural analysis of a eukaryotic sliding DNA clamp-clamp loader complex. Nature 429:724-730. - PubMed
-
- Bruning, J. B., and Y. Shamoo. 2004. Structural and thermodynamic analysis of human PCNA with peptides derived from DNA polymerase δ p66 subunit and FLAP endonuclease 1. Structure 12:2209-2219. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous