

Ubiquilin and p97/VCP bind erasin, forming a complex involved in ERAD
- PMID: 19822669
- PMCID: PMC2768832
- DOI: 10.1083/jcb.200903024
Ubiquilin and p97/VCP bind erasin, forming a complex involved in ERAD
Abstract
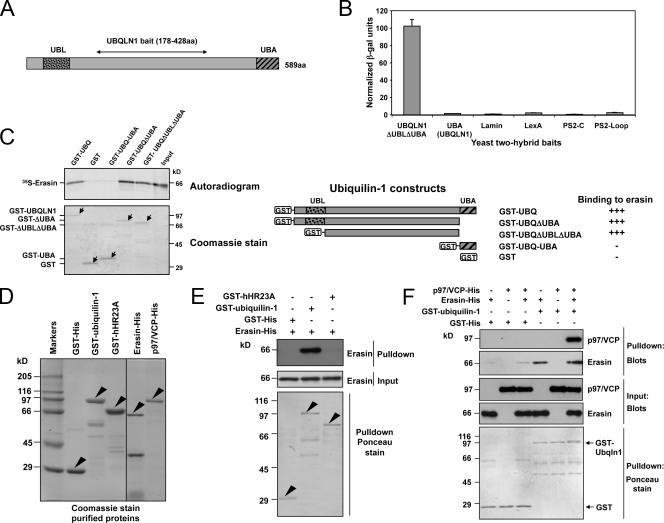
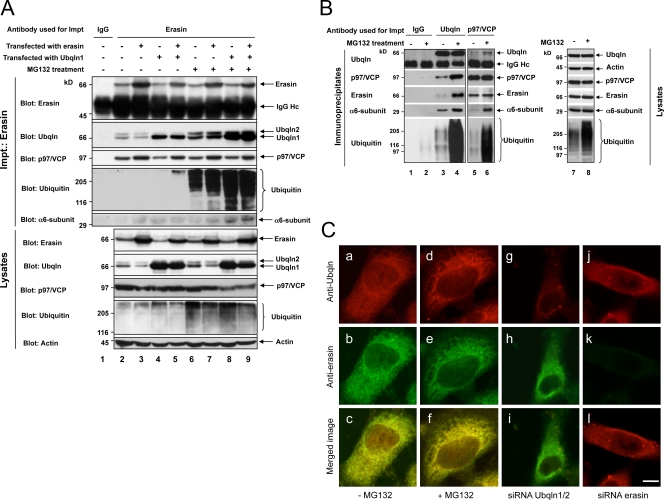
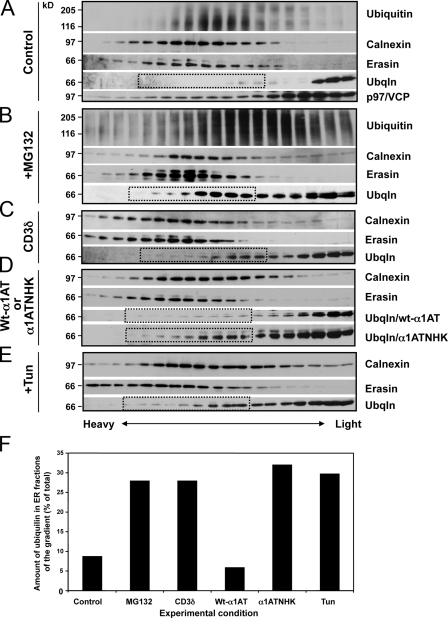
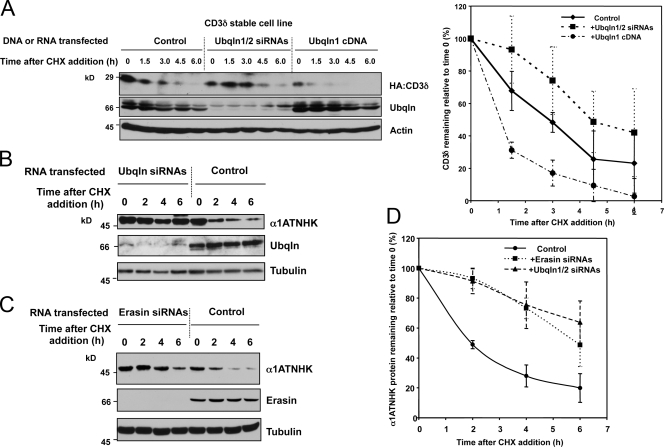
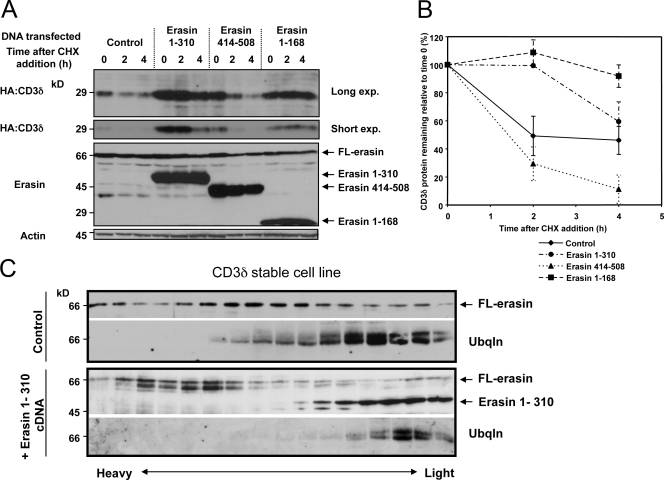
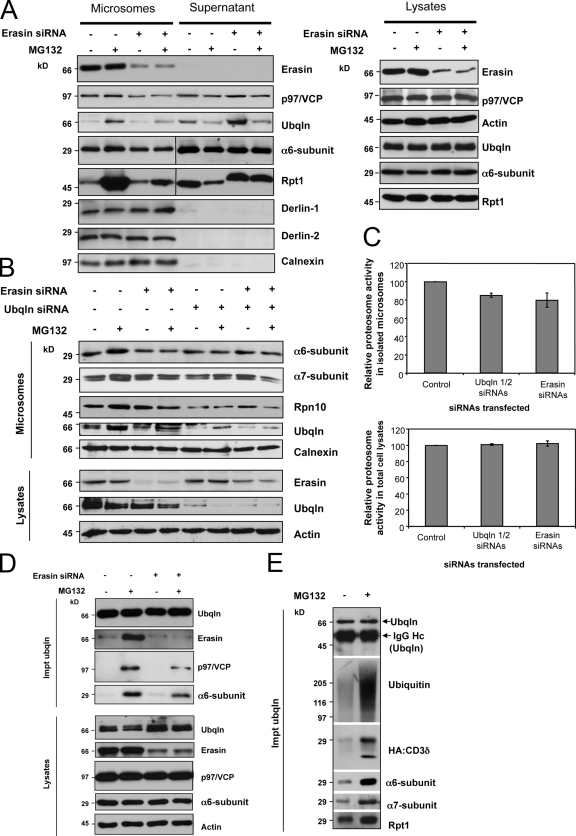
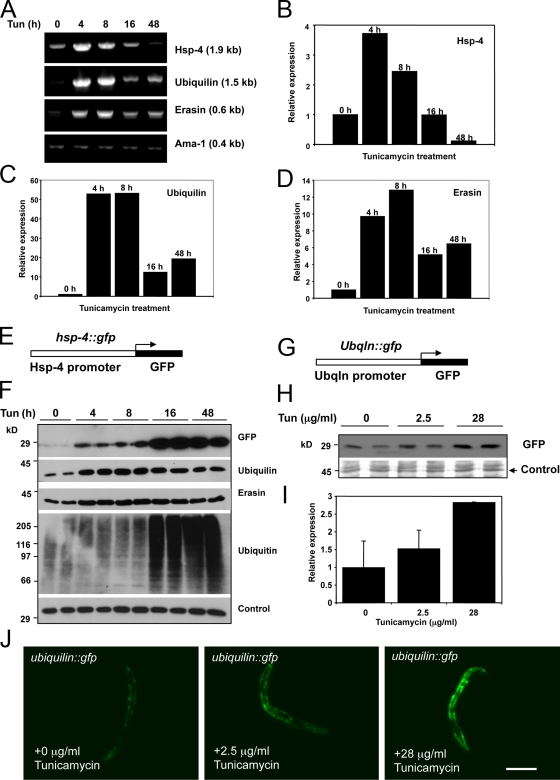
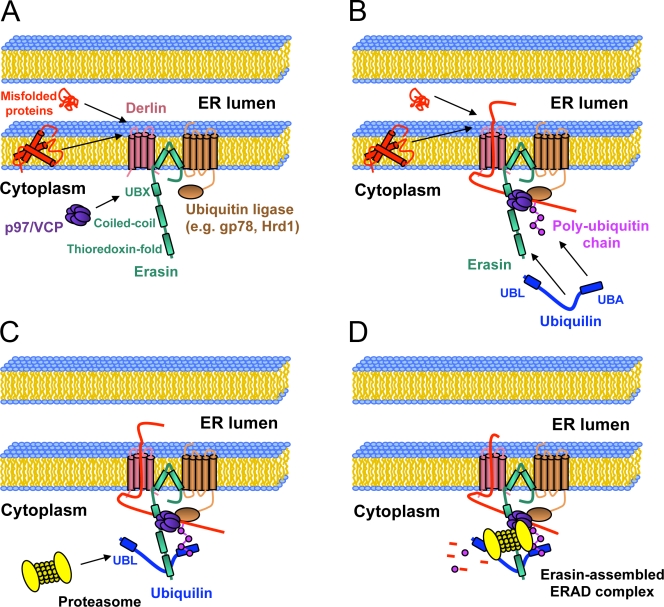
Unwanted proteins in the endoplasmic reticulum (ER) are exported into the cytoplasm and degraded by the proteasome through the ER-associated protein degradation pathway (ERAD). Disturbances in ERAD are linked to ER stress, which has been implicated in the pathogenesis of several human diseases. However, the composition and organization of ERAD complexes in human cells is still poorly understood. In this paper, we describe a trimeric complex that we propose functions in ERAD. Knockdown of erasin, a platform for p97/VCP and ubiquilin binding, or knockdown of ubiquilin in human cells slowed degradation of two classical ERAD substrates. In Caenorhabditis elegans, ubiquilin and erasin are ER stress-response genes that are regulated by the ire-1 branch of the unfolded protein response pathway. Loss of ubiquilin or erasin resulted in activation of ER stress, increased accumulation of polyubiquitinated proteins, and shortened lifespan in worms. Our results strongly support a role for this complex in ERAD and in the regulation of ER stress.
Figures


References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
