

Bioelectric controls of cell proliferation: ion channels, membrane voltage and the cell cycle
- PMID: 19823012
- PMCID: PMC2862582
- DOI: 10.4161/cc.8.21.9888
Bioelectric controls of cell proliferation: ion channels, membrane voltage and the cell cycle
Abstract
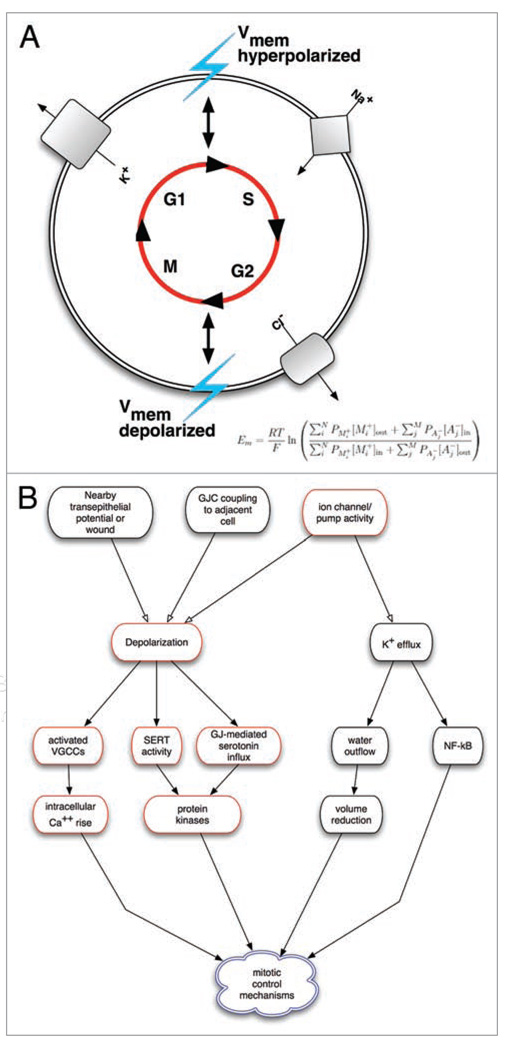
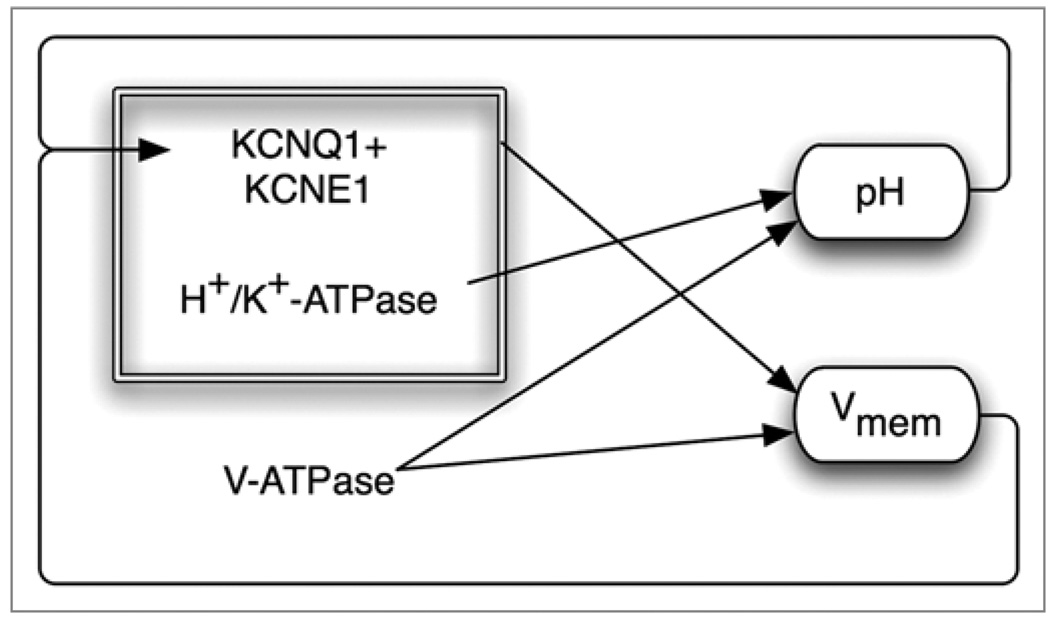
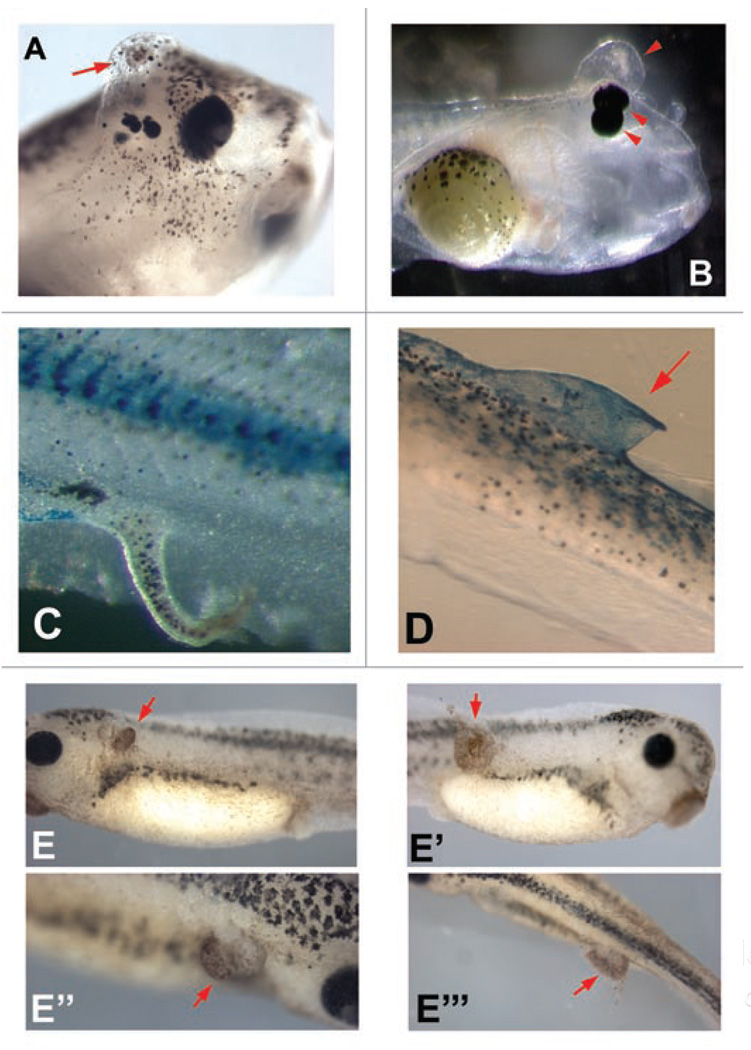
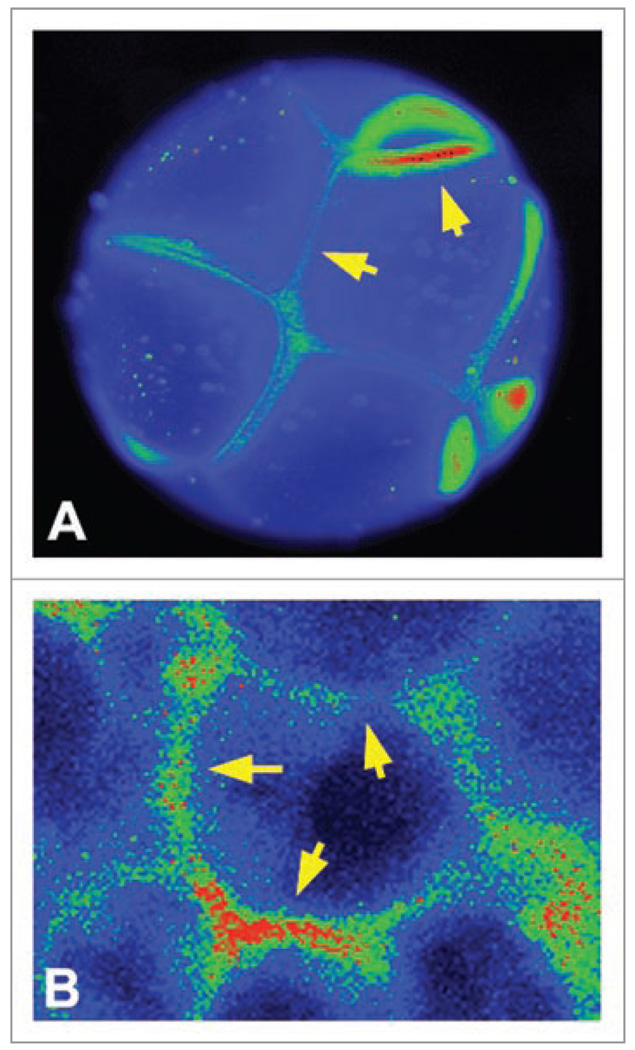
All cells possess long-term, steady-state voltage gradients across the plasma membrane. These transmembrane potentials arise from the combined activity of numerous ion channels, pumps and gap junction complexes. Increasing data from molecular physiology now reveal that the role of changes in membrane voltage controls, and is in turn controlled by, progression through the cell cycle. We review recent functional data on the regulation of mitosis by bioelectric signals, and the function of membrane voltage and specific potassium, sodium and chloride ion channels in the proliferation of embryonic, somatic and neoplastic cells. Its unique properties place this powerful, well-conserved, but still poorly-understood signaling system at the center of the coordinated cellular interactions required for complex pattern formation. Moreover, disregulation of ion channel expression and function is increasingly observed to be not only a useful marker but likely a functional element in oncogenesis. New advances in genomics and the development of in vivo biophysical techniques suggest exciting opportunities for molecular medicine, bioengineering and regenerative approaches to human health.
Figures
References
-
- Binggeli R, Weinstein R. Membrane potentials and sodium channels: hypotheses for growth regulation and cancer formation based on changes in sodium channels and gap junctions. J Theor Biol. 1986;123:377–401. - PubMed
-
- Balitsky KP, Shuba EP. Resting potential of malignant tumour cells. Acta Unio Int Contra Cancrum. 1964;20:1391–1393. - PubMed
-
- Johnstone BM. Micro-electrode penetration of ascites tumour cells. Nature. 1959;183:411. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous