

Mosquito infection responses to developing filarial worms
- PMID: 19823571
- PMCID: PMC2752998
- DOI: 10.1371/journal.pntd.0000529
Mosquito infection responses to developing filarial worms
Abstract
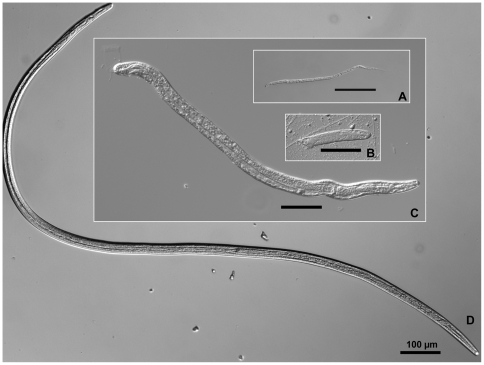
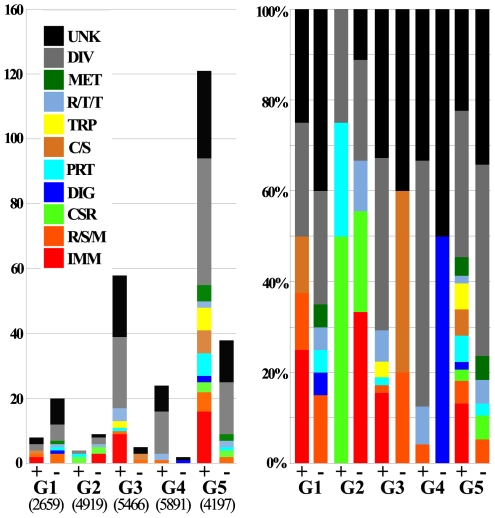
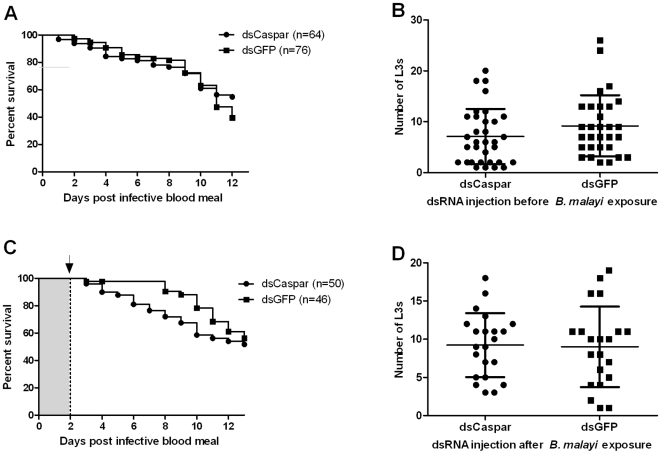
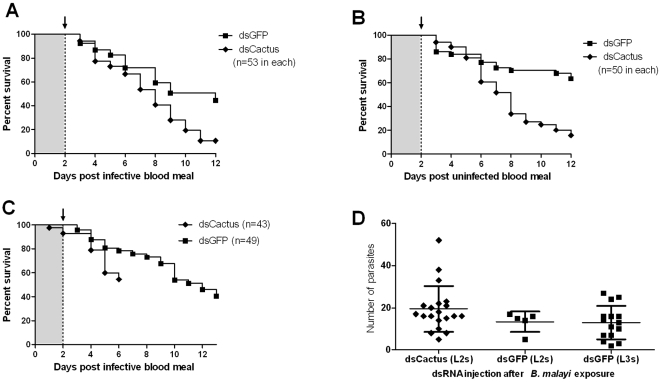
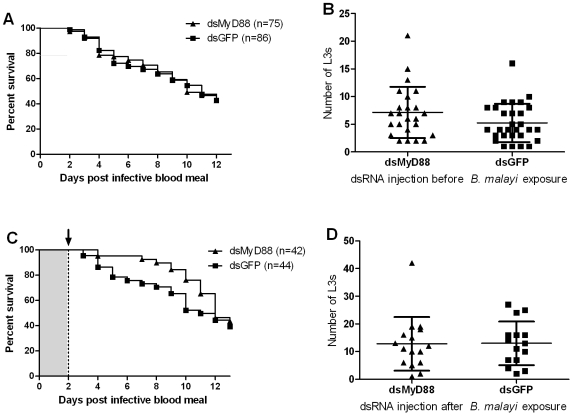
Human lymphatic filariasis is a mosquito-vectored disease caused by the nematode parasites Wuchereria bancrofti, Brugia malayi and Brugia timori. These are relatively large roundworms that can cause considerable damage in compatible mosquito vectors. In order to assess how mosquitoes respond to infection in compatible mosquito-filarial worm associations, microarray analysis was used to evaluate transcriptome changes in Aedes aegypti at various times during B. malayi development. Changes in transcript abundance in response to the different stages of B. malayi infection were diverse. At the early stages of midgut and thoracic muscle cell penetration, a greater number of genes were repressed compared to those that were induced (20 vs. 8). The non-feeding, intracellular first-stage larvae elicited few differences, with 4 transcripts showing an increased and 9 a decreased abundance relative to controls. Several cecropin transcripts increased in abundance after parasites molted to second-stage larvae. However, the greatest number of transcripts changed in abundance after larvae molted to third-stage larvae and migrated to the head and proboscis (120 induced, 38 repressed), including a large number of putative, immunity-related genes (approximately 13% of genes with predicted functions). To test whether the innate immune system of mosquitoes was capable of modulating permissiveness to the parasite, we activated the Toll and Imd pathway controlled rel family transcription factors Rel1 and Rel2 (by RNA interference knockdown of the pathway's negative regulators Cactus and Caspar) during the early stages of infection with B. malayi. The activation of either of these immune signaling pathways, or knockdown of the Toll pathway, did not affect B. malayi in Ae. aegypti. The possibility of LF parasites evading mosquito immune responses during successful development is discussed.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- WHO. Global Programme to Eliminate Lymphatic Filariasis: Annual Report on Lymphatic Filariasis 2001 . Geneva: World Health Organization.; 2002. 76p
-
- WHO. Global programme to eliminate lymphatic filariasis. Wkly Epidemiol Rec. 2007;82:361–380. - PubMed
-
- Christensen BM, Sutherland DR. Brugia pahangi: Exsheathment and midgut penetration in Aedes aegypti. Transactions of the American Microscopical Society. 1984;103:423–433.
-
- Murthy PK, Sen AB. Sequential development changes in microfilariae of subperiodic Brugia malayi to infective larvae in susceptible strain of Aedes aegypti (Macdonald). J Commun Dis. 1981;13:102–109. - PubMed
-
- Perrone JB, Spielman A. Microfilarial perforation of the midgut of a mosquito. J Parasitol. 1986;72:723–727. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
