

Inhibition of cholesterol recycling impairs cellular PrP(Sc) propagation
- PMID: 19823766
- PMCID: PMC2777232
- DOI: 10.1007/s00018-009-0158-4
Inhibition of cholesterol recycling impairs cellular PrP(Sc) propagation
Abstract
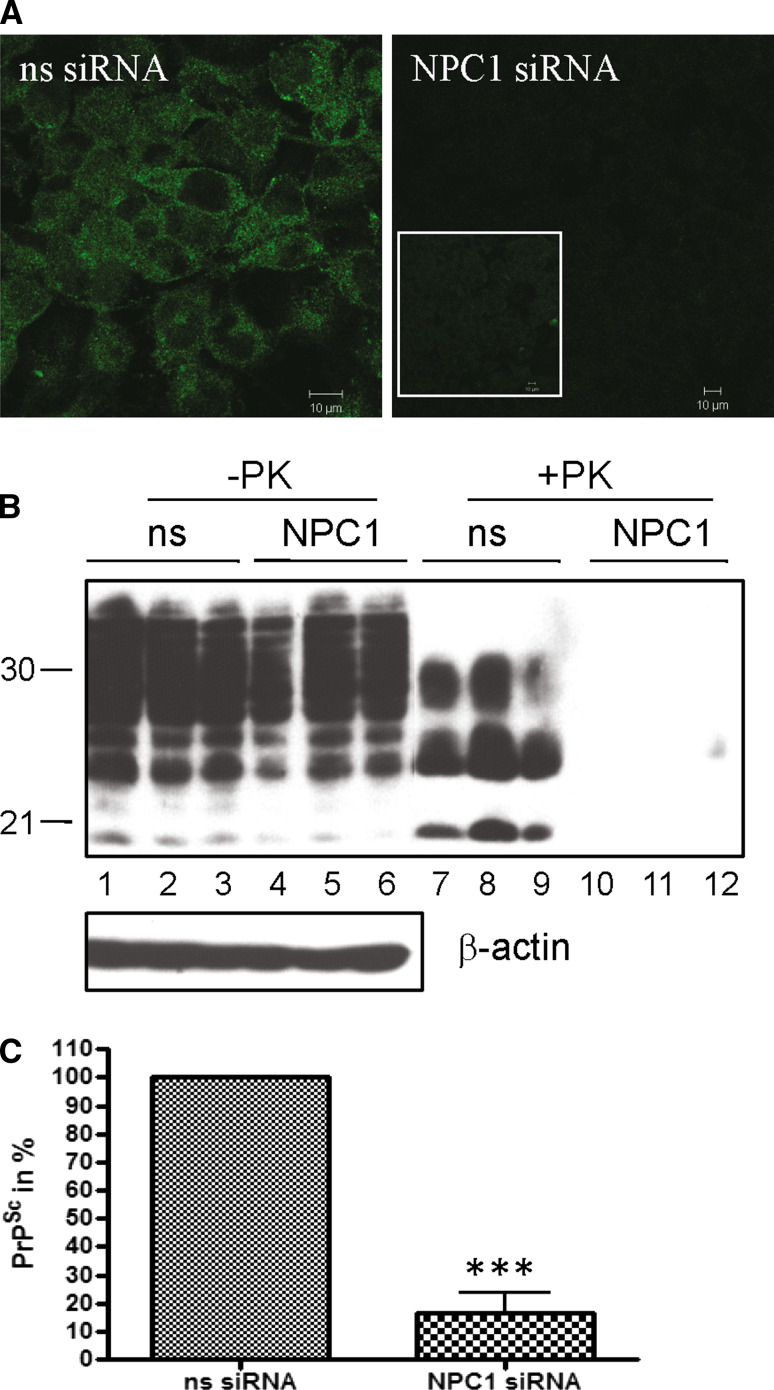
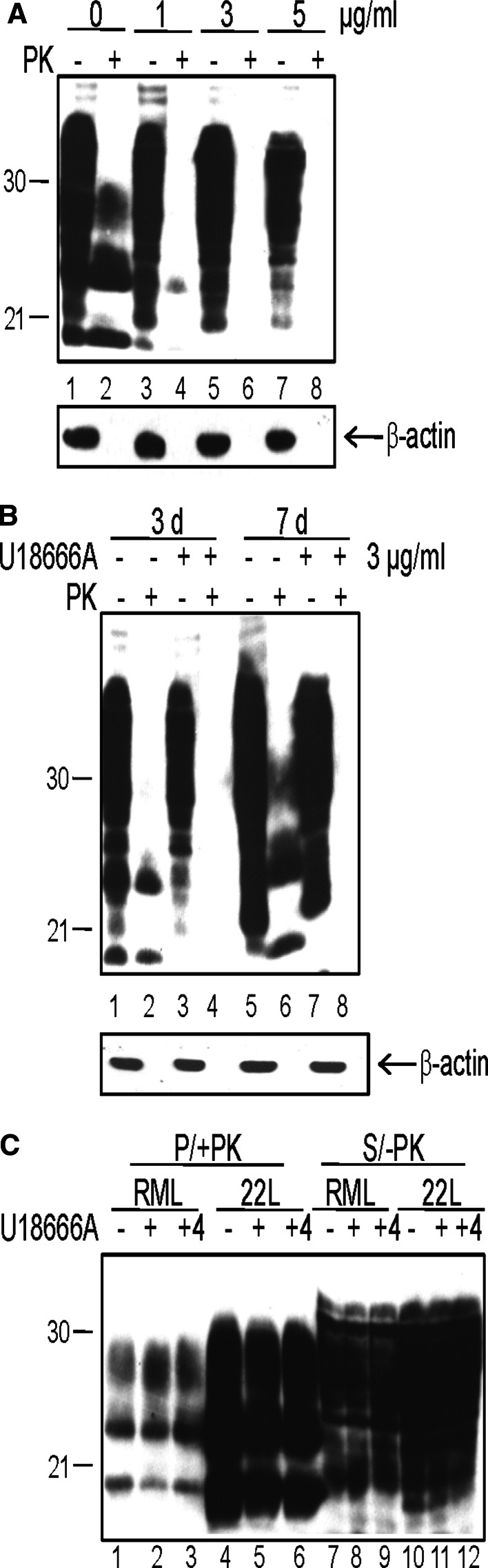
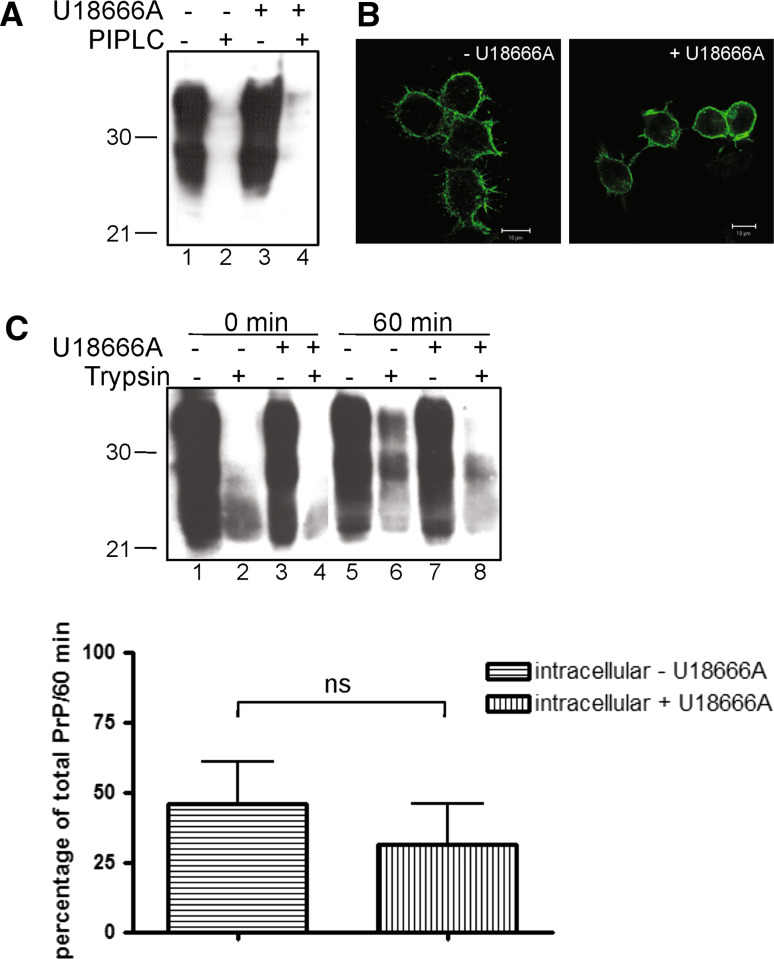
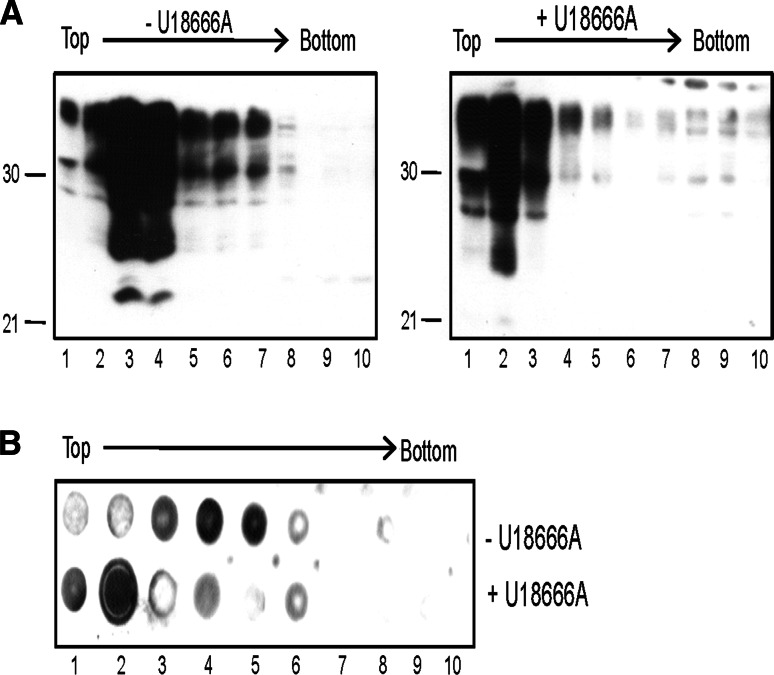
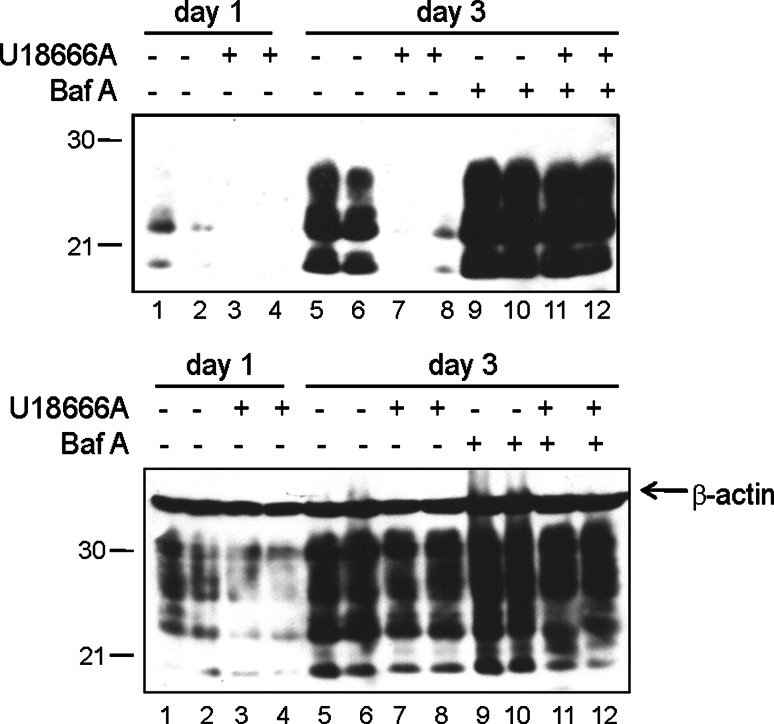
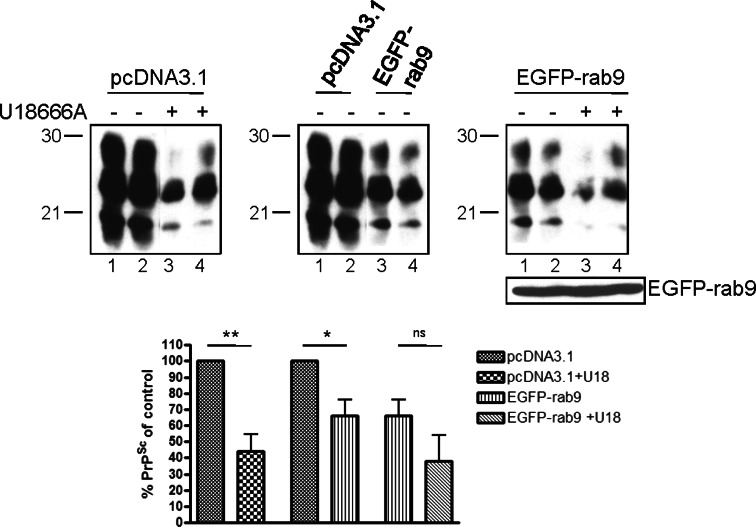
The infectious agent in prion diseases consists of an aberrantly folded isoform of the cellular prion protein (PrP(c)), termed PrP(Sc), which accumulates in brains of affected individuals. Studies on prion-infected cultured cells indicate that cellular cholesterol homeostasis influences PrP(Sc) propagation. Here, we demonstrate that the cellular PrP(Sc) content decreases upon accumulation of cholesterol in late endosomes, as induced by NPC-1 knock-down or treatment with U18666A. PrP(c) trafficking, lipid raft association, and membrane turnover are not significantly altered by such treatments. Cellular PrP(Sc) formation is not impaired, suggesting that PrP(Sc) degradation is increased by intracellular cholesterol accumulation. Interestingly, PrP(Sc) propagation in U18666A-treated cells was partially restored by overexpression of rab 9, which causes redistribution of cholesterol and possibly of PrP(Sc) to the trans-Golgi network. Surprisingly, rab 9 overexpression itself reduced cellular PrP(Sc) content, indicating that PrP(Sc) production is highly sensitive to alterations in dynamics of vesicle trafficking.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
