

Role of virus-encoded microRNAs in herpesvirus biology
- PMID: 19828316
- PMCID: PMC2802859
- DOI: 10.1016/j.tim.2009.09.002
Role of virus-encoded microRNAs in herpesvirus biology
Abstract
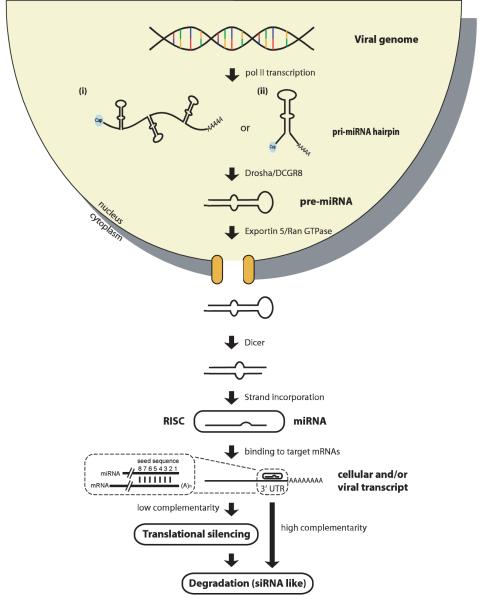
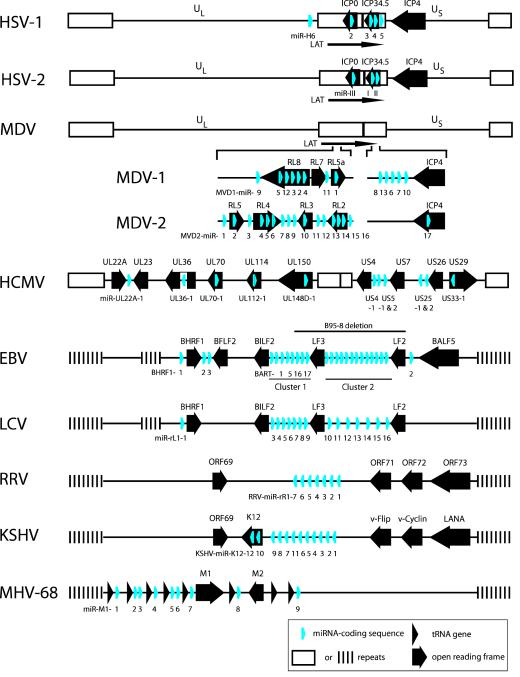
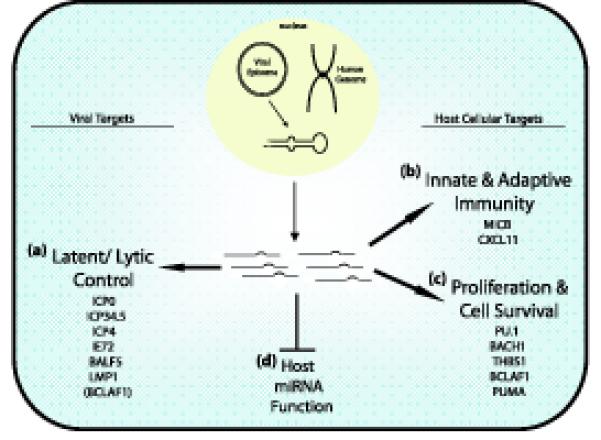
MicroRNAs (miRNAs) are short RNAs of about 22 nucleotides in length that post-transcriptionally regulate gene expression by binding to 3' untranslated regions of mRNAs, thereby inducing translational silencing. Recently, more than 140 miRNAs have been identified in the genomes of herpesviruses. Deciphering their role in viral biology requires the identification of target genes, a challenging task because miRNAs require only limited complementarity. The subject of this review will be the herpesvirus miRNAs and their respective target genes that have been determined experimentally to date. These miRNAs regulate fundamental cellular processes including immunity, angiogenesis, apoptosis, and key steps in the herpesvirus life cycle, latency and the switch from latent to lytic replication.
Figures
References
-
- Lee RC, Feinbaum RL, Ambros V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell. 1993;75(5):843–54. - PubMed
-
- Ruvkun G, et al. Dominant gain-of-function mutations that lead to misregulation of the C. elegans heterochronic gene lin-14, and the evolutionary implications of dominant mutations in pattern-formation genes. Dev Suppl. 1991;1:47–54. - PubMed
-
- Wightman B, Ha I, Ruvkun G. Posttranscriptional regulation of the heterochronic gene lin-14 by lin-4 mediates temporal pattern formation in C. elegans. Cell. 1993;75(5):855–62. - PubMed
-
- Pasquinelli AE, et al. Conservation of the sequence and temporal expression of let-7 heterochronic regulatory RNA. Nature. 2000;408(6808):86–9. - PubMed
-
- Reinhart BJ, et al. The 21-nucleotide let-7 RNA regulates developmental timing in Caenorhabditis elegans. Nature. 2000;403(6772):901–6. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
